

International Journal of Ophthalmology and Clinical Research
Ocular Hypotensive Activity of a Non-Peptide Bradykinin B2-Receptor Antagonist (WIN-64338) In Dutch-Belt Rabbits- A Case of Poly-Pharmacology in Action
Najam A Sharif*, Laura Klekar, Linya Li and Shouxi Xu
Pharmaceutical Research, Alcon Research Ltd (A Novartis Company), USA
*Corresponding author: Najam A Sharif, Global Biomedical Sciences, Santen Inc., 2100 Powell Street, Emeryville, CA 94608, USA, Tel: (415)-268-9181, E-mail: nsharif@santeninc.com
Int J Ophthalmol Clin Res, IJOCR-2-031, (Volume 2, Issue 4), Research Article; ISSN: 2378-346X
Received: April 17, 2015 | Accepted: August 07, 2015 | Published: August 11, 2015
Citation: Sharif NA, Klekar L, Li L, Xu S (2015) Ocular Hypotensive Activity of a Non-Peptide Bradykinin B2-Receptor Antagonist (WIN-64338) In Dutch-Belt Rabbits- A Case of Poly-Pharmacology in Action . Int J Ophthalmol Clin Res 2:031. 10.23937/2378-346X/1410031
Copyright: © 2015 Sharif NA, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Background: We investigated receptors and signaling mechanisms potentially mediating the intraocular pressure (IOP)-lowering due to intravitreal (ivt) injection of (S)-WIN-64338, a well-known bradykinin (BK) B2-Receptor antagonist.
Methods: Previously published ivt injection, receptor binding and signal transduction procedures were utilized.
Results: (S)-WIN-64338 lowered IOP in Dutch-belt rabbits (e.g. 47.4 ± 2.9% reduction with 50μg ivt, 72 hrs post-injection, relative to baseline and vehicle; n=7-10). Two other ivt-delivered non-peptidic B2-Receptor antagonists (and one B1-antagonist) were essentially devoid of such activity. (S)-WIN-64338 exhibited a high affinity for B2-Receptors (Ki=167 ± 47nM; n=3), and an appreciable affinity for other receptors (e.g. muscarinic, dopaminergic, serotonergic, adrenergic, neurokinin, neuropeptide Y, somatostatin and cholecystokinin), protein kinase C, Na+- and Ca2+-channels. WIN-64338 mobilized intracellular Ca2+ (13-19% at 10μM relative to BK [100% at 1μM]), and released prostaglandins (30 -163% relative to BK [100% at 1μM]) from primary human ciliary muscle and trabecular meshwork cells.
Conclusion: The long-lasting ocular hypotension induced by (S/R)-WIN-64338 may be attributed to activation/inhibition of multiple receptors and ion-channels in cells involved with aqueous humor production and/or promoting outflow.
Keywords
WIN-64338, Bradykinin, Receptors, Intraocular pressure, Glaucoma, Ocular hypertension
Introduction
Ocular hypertension is a well-recognized and treatable biomarker that is believed to be involved in the etiology of primary open-angle glaucoma, the second leading cause of blindness in the world [1-4]. This retinopathy is a painless disease that is slow but progressive and steals the patient's sight if left untreated [1-4]. Over the last several decades, a number of pharmaceutical agents have been shown to be effective in reducing intraocular pressure (IOP) and have been approved for treatment of glaucoma and the associated elevated IOP [5-8]. Thus, muscarinic agonists, beta-blockers, alpha-2 agonists, carbonic anhydrase inhibitors and FP-class prostaglandins, and some combinations thereof, are available for lowering IOP of glaucoma patients [5-10]. While these drugs have proved effective in the reduction and control of ocular hypertension via a number of mechanisms of action, they still have a variety of deficiencies including causing side-effects such as ocular surface burning, hyperemia, ocular foreign-body sensation, ocular allergy, pulmonary and cardiac insufficiency, browe-ache, and iridial and skin hyper-pigmentation [11,12]. In some instances, the drugs have a slow onset of action and a relatively short duration of action that limits the overall benefit to the patient. Due to the latter, some medications have to be dosed 2-3 times daily to keep the IOP under control and this is associated with reduced compliance and thus the drugs lose their effectiveness for glaucoma treatment [13-15]. Therefore, there continues to be an unmet medical need for more effective ocular medicines that lower and control IOP over a much longer time-frame, and have minimal or further reduced ocular and/or systemic side-effects.
In an effort to find new ocular hypotensive drug candidates, we screened a large number of compounds of various pharmacological classes in isolated human ocular cells known to be involved in mediating the effects of ocular hypotensive agents and also in various animal models of elevated IOP using different routes of administration. From such screening efforts we recently discovered the potent and effective topical ocular (t.o.) hypotensive activity of two non-peptide bradykinin (BK) B2-Receptor-selective agonists, FR-190997 [16-18] and BK2A78 [19] in ocular hypertensive eyes of cynomolgus monkeys. Additionally, we recently showed that BK (an endogenous peptide ligand of B2-Receptors) injected into the vitreous of Dutch-belted rabbits also produced robust IOP-lowering of relatively short duration [20]. In the current studies, we investigated the effect of a previously documented non-peptide BK B2-Receptor antagonist, WIN-64338 [21], on Dutch-belted rabbit IOP after topical ocular (t.o.) and intravitreal (ivt) administration. We also studied the potential receptor(s) and down-stream signal transduction pathways involved in the ocular hypotensive activity of WIN-64338 (Figure 1).
Materials and Methods
Ethical statement/institutional review and approval of studies
All animal studies reported herein were reviewed and approved by the Alcon animal studies review board which also approved all the study protocols utilized in the current studies as per Alcon procedures (e.g. PROC-002851 titled "Routes of Drug Administration"). Additional experimental guidelines were followed as described in the other Alcon procedural documents (PROC-0000123, PROC-0000071, PROC-0000067, and PROC-0002809). Guidelines for use of animals in research were also adopted as necessary per Association for Research in Vision and Ophthalmology (ARVO).
IOP measurements in dutch-belted rabbit eyes
For ivt studies, adult male Dutch-belted rabbits (n=7-10/treatment group) were anesthetized with an intramuscular injection ketamine 10mg/kg + xylazine 3mg/kg using a 1 cc syringe with a 30G1/2-inch needle. The test compounds were suspended in our standard screening vehicle formulation whose composition was as follows: 0.5% hydroxypropylmethylcellulose (HPMC, the suspending agent); 0.05% polysorbate 80 (a nonionic surfactant that wets the suspended particles); 0.2% dibasic sodium phosphate buffer (buffering agent); 0.75% NaCl (tonicity agent); 0.01% disodium edetate dihydrate (preservative aid); 0.01% benzalkolium chloride (preservative); pH adjusted to 6-7.4 using NaOH and/or HCl; 100% purified water. Freshly prepared compound solutions/suspension were utilized in each study.
For ivt studies, half of a 30G1/2-inch needle, on a Hamilton syringe loaded with 30μL vehicle or test substance suspended in the vehicle, was inserted into the vitreous, 4-5mm posterior of limbus at the superior temple quadrant of one eye of each rabbit. The needle was inserted at a 45 degree angle towards the back of the eye in order to avoid damaging the lens. The vehicle or test substance was slowly injected into the vitreous and the needle held in place for a minute after completion of the injection at which time it was slowly removed from the eye. This ensured minimal reflux of the injected liquid. IOP was determined at time-zero (after injection) and then at various time-points thereafter with an applanation pneumatonometer [17,18,20]. This intravitreal (ivt) delivery of the test agents overcame the issues of poor ocular penetration of polar drugs such as WIN-64338 (Figure 1). Two other B2-Receptor antagonists (FR-165649 and FR-173657; Figure 1) [22,23] and a B1-receptor antagonist (LF-23-1591, an analog of LF22-0542) [24] were also studied by the same ivt injection process in the Dutch-belt rabbits for comparison purposes.
In addition to the ivt administration of test agents, compounds of interest were also tested for IOP-lowering activity when instilled to conscious Dutch-belted rabbit eyes via the topical ocular (t.o.) route of administration (30μl drop) prepared in the same vehicle formulation as for ivt studies [25,26]. IOP was measured with an applanation pneumatonometer after light corneal anesthesia with 0.1% proparacaine [17,18,20,25,26]. After baseline IOP measurements were taken, one eye of each of 7-10 (or both eyes of 5) rabbits per compound dose was topically dosed with compound. Either the contralateral eye was dosed with vehicle or a separate group of rabbits was used for vehicle control. Subsequent IOP measurements were made at various intervals.
[3H]-BK and [3H]-des-Arg10-kallidin receptor binding and other ligand binding assays
In order to verify the affinity of the key test compounds of interest (used in the above studies) for the two major BK-receptor subtypes, B1-receptors and B2-Receptors, specific radio-ligand binding studies directed at these sub-types were conducted. Cell membranes of Chinese hamster ovary cells transfected with human cloned BK B1-receptors or B2-Receptors (Euroscreen [Gosselies, Belgium]; Cerep Inc. [Le Bois L'Evegue, France]; Chantest Corp., Bethesda, MD, for custom expansion of cell cultures and cell homogenates) were used in the receptor binding assays. Briefly, cell membranes were incubated with either 0.35nM [3H]-des-Arg10-kallidin [24] or 0.1-0.2nM [3H]-BK [27] (Perkin Elmer Corp., Cambridge, MA) in the absence or presence of unlabeled Arg9[Leu8]bradykinin or BK (1μM) in 96-well microtiter plates at 23℃ to radiolabel B1-receptors and B2-Receptors, respectively. Aliquots (50μL) of test agents or incubation buffer were added to various wells containing a total volume of 0.5mL incubation medium. The assay was continued for 1 hr and then the contents of the wells were harvested over GF/B glass fiber filters previously soaked in 0.5% polyethyleneimine under rapid vacuum (Tomtec Inc.; Gaithersburg, MD). The filter-bound radioactivity (Arg9[Leu8]bradykinin or [3H]-BK) was determined by liquid scintillation spectrometry and the data analyzed using a sigmoidal-fit, iterative curve-fitting computer program (ActivityBase®; IDBS, Surrey, UK) as previously described [17,20,26]. Origin Software® (Microcal Inc; Northampton, MD) was utilized to graph the data as necessary. All cumulative data were represented as mean ± SEM from the number of experiments conducted.
In order to determine the potential for (S)-WIN-64338 to bind to ligand binding sites other than those associated with the BK B2-Receptors, this compound was profiled for its ability to displace radiolabeled non-BK receptors (cell surface and intracellular), non-BK-related ligand binding sites, such as transmitter uptake sites, enzymes, ion-channels and immunological factors using 1nM, 100nM and 10μM final concentrations of (S)-WIN-64338. These studies were performed at Caliper LifeSciences [NovaScreen] (Hanover, MD) employing well document assays and procedures as previously described [28] and as can be found on the following websites for PerkinElmer, Euroscreen and Cerep: http://www.perkinelmer.com/services/contractresearch/default.xhtml; http://www.euroscreen.com; http://www.cerep.fr/Cerep/Users/index.asp. (S)-WIN-64338 was purchased from Tocris Bioscience (an R & D Systems company; Minneapolis, MN). (R)-WIN-64338 was custom synthesized for Alcon at Tocris Bioscience.
Assays to measure intracellular Ca2+ ([Ca2+]i) mobilization
Briefly, [Ca2+]i mobilization assays were conducted as previously described in detail [20,26,29] using a Fluorescence Imaging Plate Reader (FLIPR-Tetra) in conjunction with a proprietary Ca2+-sensitive dye (FLIPR Calcium Assay Kit) (Molecular Devices, Menlo Park, CA; http://www.moleculardevices.com/Products/Assay-Kits/GPCRs/FLIPR-Calcium.html). Isolated primary human ciliary muscle (h-CM) cells [30], trabecular meshwork cells (h-TM, both normal and glaucomatous) [31-34], and immortalized non-pigmented ciliary epithelial (ih-NPE) cells [35,36] of low passage number were grown to 80-90% confluency in 96-well microtiter plates (with bottom surface black). On the day of the experiment, the cells were loaded with the dye for an hour at 23℃ and the plates loaded into FLIPR. Aliquots (50μL) of test agent solutions or buffer were added to a total volume of 0.5mL/well and the change in fluorescence monitored over 2 min. BK (1μM) was used as an internal positive control agent, and the responses of the test drugs were represented as a percentage of the response induced by BK (set at 100%). When the antagonist potency was investigated, the compound (various concentrations) was added to the cells 20 min before challenging with the agonist, BK. Concentration-response/inhibition data were graphed and equilibrium inhibition constant (Ki) values calculated for each antagonist as previously described and references therein [20,27,29,34]. All data were collated and mean ± SEM determined for each compound.
Measurement of secreted prostaglandins (PGs)
Primary normal h-TM and h-CM cells isolated from human donor eyes (see above) and ih-NPE cells were distributed in 96-well microtiter plates, grown to ~95% confluency and then exposed to various concentrations of BK, buffer or test compounds. The incubation was conducted for 1 hr at 37℃ and then terminated by removal of some of the assay incubation buffer (0.5mL) which was stored at -80℃. For the determination of the total PGs released into the extracellular medium, the cell-incubation buffer was thawed slowly at 23℃ and then used in a competitive enzyme immunoassay kit (Cayman Chemical Co, Ann Arbor, MI) assay conducted per the manufacturer's instructions [17,18,20,34]. The limits of detection for the total PGs assay were 29pg/mL.
Results
Topical ocular dosing of either enantiomer of WIN-64338 failed to modulate IOP in Dutch-belted rabbits (Table 1). Similar lack of IOP-lowering efficacy of (S)-WIN-64338, and another B2-Receptor antagonist FR-165649, (both at 300μg [1%]; t.o.) was observed in ocular hypertensive monkey eyes (data not shown) out to 24 hrs post-dosing. However, a positive control agent fasudil (300μg [1%], t.o.) lowered IOP by 17-29% up to 4hr post-dose.
![]()
Table 1: Effect of topical ocular (R)-WIN-64338 on Dutch-belt rabbit IOP
View Table 1
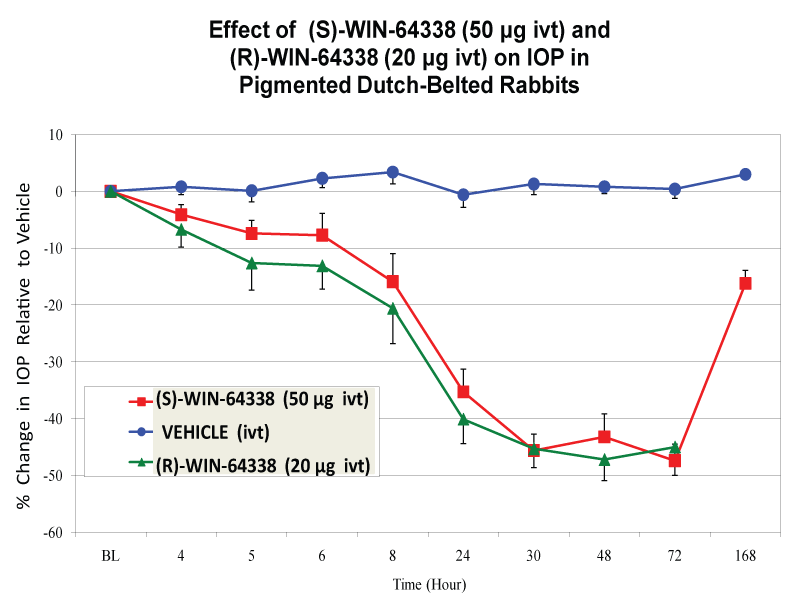
However, when (S)-or (R)-WIN-64338 (50μg [0.16%]) were delivered ivt in Dutch-belted rabbits, a pronounced IOP reduction and a long duration of action was observed with both compounds (Figure 2, Table 2). Interestingly, two other non-peptide B2-Receptor-selective antagonists, FR-165649 and FR-173657, and a non-peptide B1-receptor-selective antagonist (LF23-1591) were essentially devoid of any ocular hypotensive activity when also tested ivt in the Dutch-belted rabbits (Table 2). In an additional study, even 10μg [0.033%] of (S)-WIN-64338 ivt was quite effective in lowering IOP , thus IOP reductions of 19.5% at 8 and 24 hrs, 32.5% at 30 hrs, 36% at 48 hrs, and 27.7% at 72 hrs IVT post-dosing were observed. (R)-WIN-64338 was also quite a potent and efficacious ocular hypotensive agent (Figure 2). BK (50μg [0.16%] IVT; used as a positive control agent) reduced IOP as follows: 22.9% at 5 hrs, 37.0% at 8 hrs, and 8.3% at 24 hrs ivt post-dosing.
.
Figure 2: Changes in Dutch-belt rabbit IOPs following ivt injection of vehicle, or (R)-WIN-64338 (20μg [o.066%]) or (S)-WIN-64338 (50μg [0.16%]). After taking baseline IOP readings, one eye of each animal was injected with 30μl aliquot of each of the afore-mentioned vehicle or drugs, and the IOP measured at various times thereafter. The mean ± SEM of the IOPs from 7-10 rabbits/treatment group are shown. All observations for both enantiomers of WIN-64338 at 8 hr post-dose till 72 hr post-dose were statistically significant at levels of p<0.05-0.01, all relative to the vehicle. Note that the x-axis is not to scale but depicts the times at which the IOPs were measured after the ivt injections.
View Figure 2
![]()
Table 2: Effect of IVT administration of various non-peptide B1- and B2-Receptor antagonists on Dutch-belt rabbit IOP
View Table 2
In competition for [3H]-BK binding to human cloned B2-Receptors expressed in CHO cell membranes, (S)-WIN-64338 exhibited a moderate inhibition affinity, Ki=167 ± 47nM , Hill coefficient (nH)=1.2 ± 0.1 (n=3), and Ki=170 ± 52nM, nH=1.8 0.1 (n=3) in two separate studies. In the same assays, (R)-WIN-64338 exhibited the following [3H]-BK binding inhibition parameters: Ki=260 ± 114 nM, nH=1.3 ± 0.2 (n=3). Both compounds lacked affinity for B1-receptors labeled with [3H] Des-Arg10-Lys-BK, Ki>10μM. Profiling of (S)-WIN-64338 at 1nM-10μM against 79 other receptor/enzyme/transporter/enzyme/ion-channel binding sites revealed a significant interaction of (S)-WIN-64338 with alpha-1 and alpha-2-adrenergic, dopaminergic, histaminergic, muscarinic, opioid, orphanin, serotonergic, sigma, cholecystokinin, neurokinin, neuropeptide Y, and somatostatin receptors (Table 3). (S)-WIN-64338 also interacted relatively strongly with protein kinas C, K+-, Na+- and Ca2+-channels (Table 3).
![]()
Table 3: Ligand binding profile of (S)-WIN-64338 at numerous receptors, enzymes, transporters and ion-channels
View Table 3
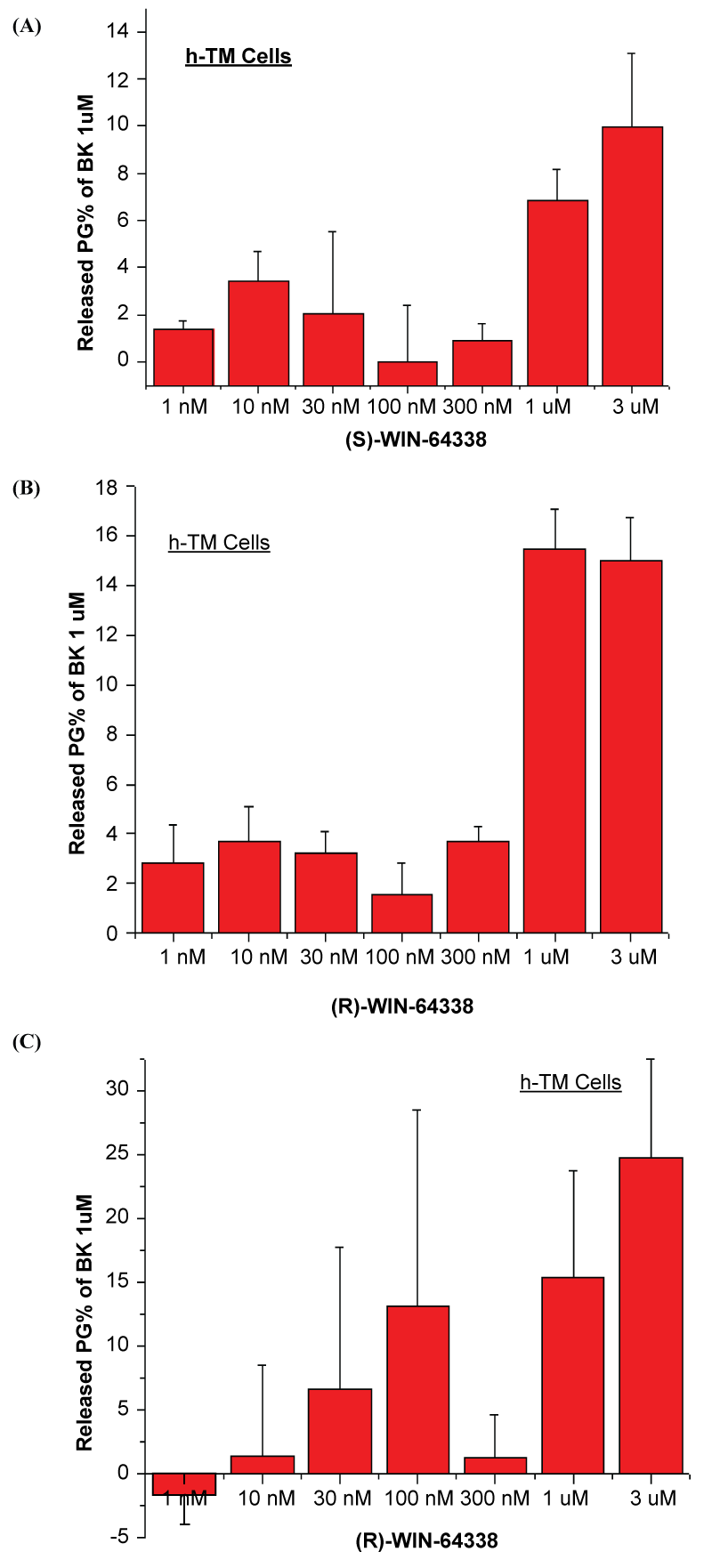
In [Ca2+]i mobilization assays using isolated primary hCM cells, (S)-WIN-64338 caused 17-19% and (R)-WIN-64338 caused 13% increase in [Ca2+]i mobilization at 10μM, behaving as weak partial agonists. Similarly, both (S)- and (R)-enantiomers of WIN-64338 (10μM) induced a small but significant amount of total PGs production and secretion from hTM cells (30 ± 1 and 31 ± 2% relative to 100% caused by 1μM BK) and from h-CM cells (126 ± 107% and 163 ± 11% relative to 100% caused by 1μM BK). Lower concentration of these enantiomers increased total PGs release from h-TM and h-CM cells to a lesser degree than BK (Figure 2A-C). For reference, 1μM BK simulated secretion of 571pg/ml total PGs from h-TM cells, while 171pg/ml total PGs were secreted from h-CM cells (detection limit was 29pg/ml).
However, when tested as an antagonist against 1μM BK in the [Ca2+]i mobilization assays under non-equilibrium conditions in normal primary isolated h-CM, h-TM cells and in ih-NPE cells, WIN-64338 exhibited relatively weak antagonist properties (Ki=157-425nM), as compared to HOE-140 (Ki=0.8-7nM) and other non-peptide B2-receoptor antagonists (Table 2).
Discussion
Recent animal studies have documented that the peptide B2-Receptor agonist, BK, but not two different the B1-receptor agonists, causes robust ocular hypotension in Dutch-belt rabbits injected ivt, and increases outflow facility in perfused bovine eye anterior chambers [20,34]. Furthermore, t.o. dosing of two different non-peptide B2-Receptor-agonist synthetic small molecules (FR-190997 and BKA278) [17,19] also induces a strong IOP-lowering response in ocular hypertensive eyes of cynomolgus monkeys that was completely blocked by a B2-Receptor-antagonist. These collective data clearly indicated that stimulation, not blockade, of the B2-Receptor is necessary for the observed reduction of IOP in the rabbits and monkeys. Thus, the remarkable magnitude and long duration of IOP-lowering induced by ivt injected enantiomers of WIN-64338 [21,37], a well-known B2-Receptor-antagonist, is unexampled to our knowledge. This apparently discrepant, yet novel, observation was quite specific for WIN-64338 since two other non-peptide B2-Receptor-antagonists (FR-165649 and FR-173657) [22,23] essentially lacked ocular hypotensive activity in the same animal model when the compounds were also delivered ivt and at the same dose as the (S)- and (R)-WIN-64-338 [21,37]. These data necessitated additional studies to help delineate the mechanism(s) involved in the WIN-64338-induced IOP reduction.
First we needed an explanation for the observation that t.o. administration of (R)-WIN-64338 in rabbits (Table 1) was ineffective in causing IOP-lowering whereas ivt injection of the same compound caused up to 50% reduction of IOP compared with the vehicle (Figure 2). An examination of the structure of WIN-64338 shows that the molecule is very polar due to the phosphonium moiety and the hydrochloride salt (Figure 1), thus rendering the drug relatively inaccessible to the anterior chamber upon t.o. dosing. Clearly, delivering the compound directly into the back of the eye by ivt injection overcomes this hurdle, although there's a delay in its action due to the diffusion needed from the vitreous to the ciliary body and trabecular meshwork (Figure 2). The fact that the (R)-enantiomer was approx. twice as potent as the (S)-enantiomer of WIN-64338 in causing ocular hypotension in the rabbits (Figure 2) indicated a stereoisomeric distinction commensurate with the slightly different functional activity of the two enantiomers. The rather slow onset of action of both enantiomers of WIN-64338 in the rabbit IOP model may be related to the fact that generation and release of other endogenous transmitters or other substances was necessary [37]. Indeed, it seems that the small amount of [Ca2+]i mobilized by WIN-64338 (see Results) followed by the induction of PG synthase and the subsequent synthesis and secretion of PGs (Figure 3A-C) are part of the reason. However, the high magnitude of the IOP-lowering response induced by both (R)- and (S)-WIN-64338 may result from recruitment of multiple receptors and their signaling pathways, including adrenergic, dopaminergic, muscarinic receptors, ion-channels, transporters, etc as revealed by the ligand binding profile of (S)-WIN-64338 (Table 3).
.
Figure 3: Total prostaglandins released by different concentrations of (S)-WIN-64338 or (R)-WIN-64338 from isolated primary, normal h-TM and h-CM (C) cells are shown. The values on the y-axes are PGs secreted (A,B) into the extracellular medium of the cells in culture plates in response to the drugs as a % of the response induced by the natural agonist of B2-Receptors, BK (1μM), the response for which was set at 100%. Only the responses induced at 1μM and 3μM of each enantiomer of WIN-64338 were statistically significant (p<0.05) relative to basal levels of PGs.
View Figure 3
The inhibition of adenosine, dopamine and serotonin transporters by (S)-WIN-64338 (Table 3) would increase the levels of these transmitters in the eye that would lead to the activation of adenosinergic [38,39], dopaminergic [40,41] and serotonergic [26,33,42] receptors that all cause ocular hypotension to different degrees mediated by various sub-types of these receptors. It is interesting to note, however, that alpha-1-antagonists [43,44], dopamine antagonists [45] and serotonin-2 (5HT2) receptor antagonists [46,47] are also potent and efficacious IOP-lowering agents in the rabbits and that there exist species differences for some of these, notably the 5HT1A and 5HT2 receptors [42,48,49]. Thus, while 5HT1A [42] agonists and 5HT2 antagonists [46,47] lower IOP in rabbits, these agents work poorly in cynomolgus monkeys [48], and actually 5HT2 agonists are robust and consistently efficacious ocular hypotensives in the latter species [33,48,49].
The opioid receptor binding activity of (S)-WIN-64338 (Table 3) may have some significance to its ability to lower IOP in the rabbits as have been demonstrated for kappa-opioid agonist bremazocine [50-52] and for mu-opioid agonist morphine [53]. These compounds modulate norepinephrine release and cause generation of nitric oxide and carbon monoxide [52,53], which then raise intracellular cGMP to cause ocular hypotension. Similarly, (S)-WIN-64338 has a relatively high affinity for sigma receptors [54], and ion-channels [54-56] for Na+, K+ and Ca2+ (Table 3). Since sigma agonists [54], and inhibitors of the latter channels have some IOP-lowering activities, it is likely that ocular hypotension caused by WIN-64338 may have some contribution from this receptor and ion-channels.
Unlike the linkage of many ligand binding sites, receptors and ion-channels to IOP reduction discussed above, the direct interaction of (S)-WIN-64338 with protein kinase C, and with receptors for cholecystokinin, neurokinin (substance P), neuropeptide Y and somatostatin (Table 3) currently does not appear have any correlative evidence available in the literature. However, since these peptides are present in ocular tissues and/or in the aqueous humor [57-60], further studies appear warranted to investigate the role of receptors of these agents in modulation of IOP.
A fascinating area connected with the multitude of affinities and functional activities of both (R)-and (S)-WIN-64338 is the role that poly-pharmacology plays in mediating reduction of elevated IOP. Pharmacological selectivity for specific receptor or enzyme targets is relative to the number of on-target and off-target activities of the compounds. Furthermore, it is the actual concentration achieved at the target protein that determines the engagement of that entity in down-stream effects of the compound in terms of activating or inhibiting the signaling pathway(s) at the cellular level. This is further complicated by the agonist/partial agonist/ inverse agonists/antagonist nature of the compounds for receptors, and activators/inhibitors for the enzymes and ion-channels. It is well known that until the discovery and use of modern pharmacological tools, delineation of receptor sub-types and enzyme sub-classes was not possible. Thus, in the current context several compounds with mixed pharmacological activities have been identified and shown to lower IOP in various animal models of ocular hypertension. For example, flesinoxan (5HT1A agonist/alpha-1 adrenoceptor-antagonist) [42], flunarizine (sigma ligand and Ca2+-channel blocker) [54,55], cabergoline (dopamine and serotonin receptor agonist) [26,40], sulprostone (EP1 and EP3 prostaglandin [PG] agonist) [60], AL-6598 (DP and EP2 PG receptor agonist) [61,62], and unoprostone (FP receptor agonist/K+-channel blocker) [63] produce robust IOP reduction probably due to their multiple pharmacological activities. Therefore, the varied and numerous receptor/transporter and ion-channel activities associated with WIN-64338 (in addition to its first described BK B2-Receptor antagonist actions; Table 3) are most likely the contributors to its profound ocular hypotensive actions in the rabbits observed in the current studies (Table 2, Figure 2). Such poly-pharmacology may be very useful since it is precisely the idea behind combination products [13,14,64,65] where differing pharmacological classes of compounds, with different mechanisms of action, are formulated together to provide fixed-dose combination ocular hypertensive medications such as dual combinations (e.g. Cosopt [timolol + dorzolamide]; Simbrinza [brinzolamide + brimonidine]; DuoTrav [Travoprost + timolol]; Combigan [bimatoprost + brimonidine]) or triple combinations (e.g. dorzolamide + brimonidine + timolol) [13,14,64,65].
In addition to the above poly-pharmacology, there's a distinct possibility that depending on the cells, tissues, and animal species being utilized in the in vitro and in vivo studies, WIN-64338 may exhibit partial agonist or antagonist properties. Thus, in the current studies we observed a small (13-19% above baseline) partial agonist effect of (S)-WIN-64338 on [Ca2+]i mobilization in human ocular cells involved in aqueous humor regulation (h-CM and h-TM cells). Similarly, a 30% and 126-163% increase in PGs release from h-TM and h-CM cells, respectively, was observed when exposed to WIN-64338. Such mixed pharmacological actions of WIN-64338 are not unprecedented since another BK B2-Receptor antagonist, HOE-140, has behaved as an agonist and antagonist in isolated blood vessels of different species [66], and is a potent mitogenic agonist in a number of cell-lines [67], despite displaying classical antagonist activity in numerous other biological systems [68-70]. Moreover, there's the possibility that WIN-64338 is an activating ligand for a new sub-type of BK receptor, as has been proposed from functional studies conducted in other tissues and species [71-75].
The relatively small increase in [Ca2+]i mobilization and PGs secretion induced by (S/R)-WIN-64338 may be considered inadequate to affect IOP. However, due to signal amplification and recruitment of additional down-stream signal transduction processes associated with receptor activation, especially PG receptors [76] that involves matrix met alloproteinase (MMP) release [77], the amount of PGs released by (S/R)-WIN-64338 are more than sufficient to trigger various PG receptors in target cells involved in AQH regulation [20,32,33,36]. Thus, using the data from figure 3A-3C (also see Results), it becomes apparent that upon secretion h-TM and h-CM cells into the extracellular medium, concentrations of 0.15-0.21nM PGs can be achieved. Since PGs are very potent agents with half-maximal receptor activation of EP3 receptors at 0.41-0.63nM (by PGE2 and PGF2α) [78], and of EP4 receptors at 0.17-0.22nM (by PGE1 and PGE2) [78-80], for instance, appreciable receptor activation would be expected to occur that would result in generation of MMPs [77] that in turn degrade extracellular matrix to lower IOP.
Conclusions
In conclusion, the effective IOP-lowering activity of both enantiomers of WIN-64338 observed in Dutch-belt rabbits after ivt injections is indicated to be due to the poly-pharmacology and mixed agonist/antagonist properties associated with this compound. Discovery of other compounds that may provide such robust ocular hypotension is eagerly awaited.
References
-
Quigley HA (1996) Number of people with glaucoma worldwide. Br J Ophthalmol 80: 389-393.
-
Congdon N, O'Colmain B, Klaver CC, Klein R, Muñoz B, et al. (2004) Causes and prevalence of visual impairment among adults in the United States. Arch Ophthalmol 122: 477-485.
-
Quigley HA, Broman AT (2006) The number of people with glaucoma worldwide in 2010 and 2020. Br J Ophthalmol 90: 262-267.
-
(2000) The Advanced Glaucoma Intervention Study (AGIS): 7. The relationship between control of intraocular pressure and visual field deterioration.The AGIS Investigators. Am J Ophthalmol 130: 429-440.
-
Kersey T, Clement CI, Bloom P, Cordeiro MF (2013) New trends in glaucoma risk, diagnosis & management. Indian J Med Res 137: 659-668.
-
Sharif NA, Klimko PG (2007) CNS: Ophthalmic Agents. In: Taylor JB, Triggle DJ (ed) Comprehensive Medicinal Chemistry II., Vol. 6, Chapter 12, Elsevier, Oxford 297-320.
-
Chen J, Runyan SA, Robinson MR (2011) Novel ocular antihypertensive compounds in clinical trials. Clin Ophthalmol 5: 667-677.
-
Beidoe G, Mousa SA (2012) Current primary open-angle glaucoma treatments and future directions. Clin Ophthalmol 6: 1699-1707.
-
Bucolo C, Salomone S, Drago F, Reibaldi M, Longo A, et al. (2013) Pharmacological management of ocular hypertension: current approaches and future prospective. Curr Opin Pharmacol 13: 50-55.
-
Kaufman PL, Rasmussen CA (2012) Advances in glaucoma treatment and management: outflow drugs. Invest Ophthalmol Vis Sci 53: 2495-2500.
-
Osborne SA, Montgomery DM, Morris D, McKay IC (2005) Alphagan allergy may increase the propensity for multiple eye-drop allergy. Eye (Lond) 19: 129-137.
-
Baudouin C, Pisella PJ, Fillacier K, Goldschild M, Becquet F, et al. (1999) Ocular surface inflammatory changes induced by topical antiglaucoma drugs: human and animal studies. Ophthalmology 106: 556-563.
-
Holló G, Topouzis F, Fechtner RD (2014) Fixed-combination intraocular pressure-lowering therapy for glaucoma and ocular hypertension: advantages in clinical practice. Expert Opin Pharmacother 15: 1737-1747.
-
Baiza-Durán LM, Alvarez-Delgado J, Contreras-Rubio AY, Medrano-Palafox J, De Luca-Brown A, et al. (2009) The efficacy and safety of two fixed combinations: timolol-dorzolamide-brimonidine versus timolol-dorzolamide. A prospective, randomized, double-masked, multi-center, 6-month clinical trial. Ann Ophthalmol (Skokie) 41: 174-178.
-
Chew SK, Skalicky SE, Goldberg I (2014) Brinzolamide plus brimonidine for the treatment of glaucoma: an update. Expert Opin Pharmacother 15: 2461-2471.
-
Sharif NA, Katoli P, Kelly C, Li L, Xu S, et al. (2013) Peptide and non-peptide bradykinin (BK) agonists and antagonists help define functional BK B2-Receptors in human trabecular meshwork and ciliary body. ARVO Abst # 1997.
-
Sharif NA, Katoli P, Scott D, Li L, Kelly C, et al. (2014) FR-190997, a non-peptide bradykinin B2-Receptor partial agonist, is a potent and efficacious intraocular pressure lowering agent in ocular hypertensive cynomolgus monkeys. Drug Develop Res 75: 211-223.
-
Sharif NA, Li L, Katoli P, Xu S, Veltman J, et al. (2014) Preclinical pharmacology, ocular tolerability and ocular hypotensive efficacy of a novel non-peptide bradykinin mimetic small molecule. Exp Eye Res 128: 170-180.
-
Prasanna G, Sharif N, Li B, Hellberg M, Krause T, et al. (2014) BK2A78: a novel non-peptide bradykinin B2 agonist lowers intraocular pressure (IOP) in ocular hypertensive cynomolgus monkeys. Invest Ophthalmol Vis Sci 55: 2883.
-
Sharif NA, Xu S, Li L, Katoli P, Kelly CR, et al. (2013) Protein expression, biochemical pharmacology of signal transduction, and relation to intraocular pressure modulation by bradykinin B2 receptors in ciliary muscle. Mol Vis 19: 1356-1370.
-
Sawutz DG, Salvino JM, Dolle RE, Casiano F, Ward SJ, et al. (1994) The nonpeptide WIN 64338 is a bradykinin B2 receptor antagonist. Proc Natl Acad Sci U S A 91: 4693-4697.
-
Asano M, Hatori C, Sawai H, Johki S, Inamura N, et al. (1998) Pharmacological characterization of a nonpeptide bradykinin B2 receptor antagonist, FR165649, and agonist, FR190997. Br J Pharmacol 124: 441-446.
-
Asano M, Inamura N, Hatori C, Sawai H, Fujiwara T, et al. (1997) The identification of an orally active, nonpeptide bradykinin B2 receptor antagonist, FR173657. Br J Pharmacol 120: 617-624.
-
Porrreca F, Vanderah TW, Guo W, Barth M, dodey P, Peyrou V, et al. (2006) Antinociceptive pharmacology of N-[[4-(4,5-dihydro-1H-imidazol-2-yl)phenyl]methy]-2[2-[[(4-methoxy-2,6-dimethylphenyl) sulfonyl]methylamino]ethhoxy] -N-methylacetamide, fumerate (LF22-0542), a novel nonpeptiidic bradykinin B1 receptor antagonist. J Pharmacol Exp Ther 318: 195-205.
-
Hellberg MR, McLaughlin MA, Sharif NA, DeSantis L, Dean TR, et al. (2002) Identification and characterization of the ocular hypotensive efficacy of travoprost, a potent and selective FP prostaglandin receptor agonist, and AL-6598, a DP prostaglandin receptor agonist. Surv Ophthalmol 47: S13-33.
-
Sharif NA, McLaughlin MA, Kelly CR, Katoli P, Drace C, et al. (2009) Cabergoline: Pharmacology, ocular hypotensive studies in multiple species, and aqueous humor dynamic modulation in the Cynomolgus monkey eyes. Exp Eye Res 88: 386-397.
-
Sharif NA, Whiting RL (1993) The neuropeptide bradykinin stimulates phosphoinositide turnover in HSDM1C1 cells: B2 antagonist sensitive responses and receptor binding studies. Neurochem Res 12: 1313-1320.
-
Sweetnam PM, Lancaster J, Snowman A, Collins JL, Perschke S, et al. (1995) Receptor binding profile suggests multiple mechanisms of action are responsible for ibogaine's putative anti-addictive activity. Psychopharmacology (Berl) 118: 369-376.
-
Kelly CR, Williams GW, Sharif NA (2003) Real-time intracellular Ca2+ mobilization by travoprost acid, bimatoprost, unoprostone, and other analogs via endogenous mouse, rat, and cloned human FP prostaglandin receptors. J Pharmacol Exp Ther 304: 238-245.
-
Husain S, Farahdiba J, Crosson CE (2005) Acute effects of PGF2a on MMP-2 secretion from human ciliary muscle cells: A PKC- and ERK-dependent process. Invest Ophthalmol Vis Sci 46: 1706-1713.
-
Liu X, Lambert W, Agarwal R, Talati M, Cross W, et al. (2001) Human trabecular meshwork cells express the ciliary neurotrophic factor (cntf) tripartite receptor complex. Exp Eye Res 72: 711-717.
-
Sharif NA, Kelly CR, Crider JY (2003) Human trabecular meshwork cell responses induced by bimatoprost, travoprost, unoprostone, and other FP prostaglandin receptor agonist analogues. Invest Ophthalmol Vis Sci 44: 715-721.
-
Sharif NA, Kelly CR, McLaughlin M (2006) Human trabecular meshwork cells express functional serotonin-2A (5HT2A) receptors: role in IOP reduction. Invest Ophthalmol Vis Sci 47: 4001-4010.
-
Sharif NA, Katoli P, Kelly CR, Li L, Xu S, Wang Y, Klekar L, Earnest D, Yacoub S, Hamilton G, Jacobson N, Shepard AR, Ellis D. (2014) Trabecular meshwork bradykinin receptors: mRNA levels, immunohistochemical visualization, signaling processes pharmacology and linkage to IOP Reduction. J Ocular Pharmacol Ther 30: 21-34.
-
Coca-Prados M, Wax MB (1986) Transformation of human ciliary epithelial cells by simian virus 40: induction of cell proliferation and retention of beta 2-adrenergic receptors. Proc Natl Acad Sci U S A 83: 8754-8758.
-
Sharif NA, Wang Y, Katoli P, Xu S, Kelly CR, et al. (2014) Human non-pigmented ciliary epithelium bradykinin B2-Receptors: receptor localization, pharmacological characterization of intracellular Ca2+ mobilization, and prostaglandin secretion. Curr Eye Res. 39: 378-389.
-
Sawutz DG, Salvino JM, Dolle RE, Seone PR, Farmer SG. (1995) Pharmacology and structure-activity relationships of the nonpeptide bradykinin receptor antagonist WIN-64338. Can J Physiol Pharmacol 73: 805-8011.
-
Crosson CE, Sloan CF, Yates PW (2005) Modulation of conventional outflow facility by the adenosine A1 agonist N6-cyclohexyladenosine. Invest Ophthalmol Vis Sci 46: 3795-3799.
-
Zhong Y, Yang Z, Huang WC, Luo X (2013) Adenosine, adenosine receptors and glaucoma: an updated overview. Biochim Biophys Acta 1830: 2882-2890.
-
Platania CB, Leggio GM, Drago F, Salomone S, Bucolo C (2013) Regulation of intraocular pressure in mice: structural analysis of dopaminergic and serotonergic systems in response to cabergoline. Biochem Pharmacol 86: 1347-1356.
-
Ogidigben M, Chu TC, Potter DE (1993) Ocular hypotensive action of a dopaminergic (DA2) agonist, 2,10,11-trihydroxy-N-n-propylnoraporphine. J Pharmacol Exp Ther 267: 822-827.
-
Chidlow G, Cupido A, Melena J, Osborne NN (2001) Flesinoxan, a 5-HT1A receptor agonist/alpha 1-adrenoceptor antagonist, lowers intraocular pressure in NZW rabbits. Curr Eye Res 23: 144-153.
-
Zhan GL, Toris CB, Camras CB, Wang YL, Yablonski ME (1998) Bunazosin reduces intraocular pressure in rabbits by increasing uveoscleral outflow. J Ocul Pharmacol Ther 14: 217-228.
-
Mittag TW, Tormay A, Severin C, Podos SM (1985) Alpha-adrenergic antagonists: correlation of the effect on intraocular pressure and on alpha 2-adrenergic receptor binding specificity in the rabbit eye. Exp Eye Res 40: 591-599.
-
Chiou GC (1984) Treatment of ocular hypertension and glaucoma with dopamine antagonists. Ophthalmic Res 16: 129-134.
-
Chiou GC, Li BH (1992) Ocular hypotensive actions of serotonin antagonist-ketanserin analogs. J Ocul Pharmacol 8: 11-21.
-
Tekat D, Guler C, Arici M, Topalkara A, Erdogan H (2001) Effect of ketanserin administration on intraocular pressure. Ophthalmologica 215: 419-423.
-
May JA, McLaughlin MA, Sharif NA, Hellberg MR, Dean TR (2003) Evaluation of the ocular hypotensive response of serotonin 5-HT1A and 5-HT2 receptor ligands in conscious ocular hypertensive cynomolgus monkeys. J Pharmacol Exp Ther 306: 301-309.
-
Sharif NA, McLaughlin MA, Kelly CR (2007) AL-34662: a potent, selective, and efficacious ocular hypotensive serotonin-2 receptor agonist. J Ocul Pharmacol Ther 23: 1-13.
-
Moore TT, Potter DE (2001) Kappa opioid agonist-induced changes in IOP: correlation with 3H-NE release and cAMP accumulation. Exp Eye Res 73: 167-178.
-
Rasmussen CA, Gabelt BT, Kaufman PL (2007) Aqueous humor dynamics in monkeys in response to the kappa opioid agonist bremazocine. Trans Am Ophthalmol Soc 105: 225-238.
-
Russell-Randall KR, Dortch-Carnes J (2011) Kappa opioid receptor localization and coupling to nitric oxide production in cells of the anterior chamber. Invest Ophthalmol Vis Sci 52: 5233-5239.
-
Stagni E, Bucolo C, Motterlini R, Drago F (2010) Morphine-induced ocular hypotension is modulated by nitric oxide and carbon monoxide: role of mu3 receptors. J Ocul Pharmacol Ther 26: 31-35.
-
Wang RF, Gagliuso DJ, Podos SM (2008) Effect of flunarizine, a calcium channel blocker, on intraocular pressure and aqueous humor dynamics in monkeys. J Glaucoma 17: 73-78.
-
Campana G, Bucolo C, Murari G, Spampinato S (2002) Ocular hypotensive action of topical flunarizine in the rabbit: role of sigma 1 recognition sites. J Pharmacol Exp Ther 303: 1086-1094.
-
Jacob TJ, Civan MM (1996) Role of ion channels in aqueous humor formation. Am J Physiol 271: C703-720.
-
Stone RA, Kuwayama Y, Laties AM (1987) Regulatory peptides in the eye. Experientia 43: 791-800.
-
Kuwayama Y, Stone RA (1987) Distinct substance P and calcitonin gene-related peptide immunoreactive nerves in the guinea pig eye. Invest Ophthalmol Vis Sci 28: 1947-1954.
-
Rittig MG, Licht K, Funk RH (1993) Innervation of the ciliary process vasculature and epithelium by nerve fibers containing catecholamines and neuropeptide Y. Ophthalmic Res 25: 108-118.
-
Ghosh S, Choritz L, Geibel J, Coca-Prados M (2006) Somatostatin modulates PI3K-Akt, eNOS and NHE activity in the ciliary epithelium. Mol Cell Endocrinol 253: 63-75.
-
Waterbury LD, Eglen RM, Faurot GF, Cooper GF (1990) EP3, but not EP2, FP, or TP prostanoid-receptor stimulation may reduce intraocular pressure. Invest Ophthalmol Vis Sci 31: 2560-2567.
-
Sharif NA, Williams GW, Crider JY, Xu SX, Davis TL (2004) Molecular pharmacology of the DP/EP2 class prostaglandin AL-6598 and quantitative autoradiographic visualization of DP and EP2 receptor sites in human eyes. J Ocul Pharmacol Ther 20: 489-508.
-
Cuppoletti J, Malinowska DH, Tewari KP, Chakrabarti J, Ueno R (2012) Unoprostone isopropyl and metabolite M activate BK channels and prevent ET-1-induced [Ca2+]i increases in human trabecular meshwork and smooth muscle. Invest Ophthalmol Vis Sci 53: 5178-5189.
-
Khouri AS, Realini T, Fechtner RD (2007) Use of fixed-dose combination drugs for the treatment of glaucoma. Drugs Aging 24: 1007-1016.
-
Higginbotham EJ (2010) Considerations in glaucoma therapy: fixed combinations versus their component medications. Clin Ophthalmol 4: 1-9.
-
Félétou M, Germain M, Thurieau C, Fauchère JL, Canet E (1994) Agonistic and antagonistic properties of the bradykinin B2 receptor antagonist, Hoe 140, in isolated blood vessels from different species. Br J Pharmacol 112: 683-689.
-
Drube S, Liebmann C (2000) In various tumour cell lines the peptide bradykinin B(2) receptor antagonist, Hoe 140 (Icatibant), may act as mitogenic agonist. Br J Pharmacol 131: 1553-1560.
-
Wiernas TK, Griffin BW, Sharif NA (1997) The expression of functionally-coupled B2-bradykinin receptors in human corneal epithelial cells and their pharmacological characterization with agonists and antagonists. Br J Pharmacol 121: 649-656.
-
Heitsch H (2002) Non-peptide antagonists and agonists of the bradykinin B(2) receptor. Curr Med Chem 9: 913-928.
-
Leeb-lundberg, LM, Marceau F, Muller-Esteri W, Pettibone DJ, Zuraw BL (2005) International union of pharmacology. XLV. Classification of the kinin receptor family: from molecular mechanisms to pathophysiological consequences. Pharmacol Rev 57: 27-77.
-
Farmer SG, Burch RM, Meeker SA, Wilkins DE (1989) Evidence for a pulmonary B3 bradykinin receptor. Mol Pharmacol 36: 1-8.
-
Regoli D, Gobeil F, Nguyen QT, Jukic D, Seoane PR, et al. (1994) Bradykinin receptor types and B2 subtypes. Life Sci 55: 735-749.
-
Fathy DB, Leeb T, Mathis SA, Leeb-Lundberg LM (1999) Spontaneous human B2 bradykinin receptor activity determines the action of partial agonists as agonists or inverse agonists. Effect of basal desensitization. J Biol Chem 274: 29603-29606.
-
Cassano G, Susca F, Lippe C, Guanti G (1999) Two B1 and B2 bradykinin receptor antagonists fail to inhibit the Ca2+ response elicited by bradykinin in human skin fibroblasts. Gen Pharmacol 32: 239-244.
-
Griesbacher T, Rainer I (1999) 5-hydroxytryptamine release from skin mast cells in vivo induced by peptide but not by nonpeptide ligands for bradykinin receptors. Immunopharmacology 43: 195-201.
-
Kaplan-Messas A, Naveh N, Avni I, Marshall J (2003) Ocular hypotensive effects of cholinergic and adrenergic drugs may be influenced by prostaglandins E2 in the human and rabbit eye. Eur J Ophthalmol 13: 18-23.
-
Weinreb RN, Kashiwagi K, Kashiwagi F, Tsukahara S, Lindsey JD (1997) Prostaglandins increase matrix met alloproteinase release from human ciliary smooth muscle cells. Invest Ophthalmol Vis Sci 38: 2772-2780.
-
Abramovitz M, Adam M, Boie Y, Carrière M, Denis D, et al. (2000) The utilization of recombinant prostanoid receptors to determine the affinities and selectivities of prostaglandins and related analogs. Biochim Biophys Acta 1483: 285-293.
-
Woodward DF, Nilsson SF, Toris CB, Kharlamb AB, Nieves AL, et al. (2009) Prostanoid EP4 receptor stimulation produces ocular hypotension by a mechanism that does not appear to involve uveoscleral outflow. Invest Ophthalmol Vis Sci 50: 3320-3328.
-
Sharif NA (2002) Non-Peptide Bradykinin Receptor Antagonists for Use in Controlling Intraocular Pressure and Treating Glaucoma. US patent 6,500,831.
