Introduction
The Pfizer and Moderna COVID-19 vaccines are composed of lipid nanoparticles (LNP) containing a modified messenger RNA (mRNA) that encodes for the Spike S protein [1-3]. The LNP transfection likely involves particle engulfment by the host immune cells due to their resemblance to apoptotic bodies, vesicles with externalized phosphatidylserine (ePS). As the LNPs are decorated with PS-like ionizable phospholipids, including 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC), they encourage human phagocytes into internalizing them [4-6].
The LNP technology, to put it simply, mimics viral envelopes with ePS, a universal "eat me" signal, that directs immune cells to engulf the particle [7,8]. However, as ePS is also a potential "fuse me" signal, LNP may inadvertently facilitate the formation of pathological syncytia [9,10]. Moreover, ePS may activate a disintegrin and metalloprotease 10 and 17 (ADAM10) (ADAM 17), master regulators of syncytia formation, contributing further to the unintended consequence of cell-cell fusion [11,12].
LNP-incorporated mRNA comprises an enormous technological success that goes beyond vaccines, opening new avenues for developing "smart" therapeutics that can be delivered with pinpoint precision to specific subcellular structures [13]. The development of such therapeutics is anticipated to redefine clinical pathways, including for non-communicable diseases. However, are these therapies ready for worldwide application in their present molecular form?
The question has been asked before, often in relation to the potential toxicity of lipid formulations used in the past, especially as part of the delivering cancer therapeutics [14,15]. However, in the following sections, we take a closer look at a novel perspective, namely the mRNA vaccines' structure and composition and at their unintended biological consequences derived from pathological cell-cell fusion.
Messenger RNA Vaccines, an Overview
To elicit the formation of neutralizing antibodies, exogenously administered mRNA must avoid two key obstacles: Hydrolysis by extracellular RNAases and recognition by cytosolic innate immune sensors, including toll-like receptors (TLRs) and retinoic acid-inducible gene I (RIG-I) protein [16,17]. The former is accomplished by hiding the nucleic acid backbone into LNPs, while the latter by attaching nucleobases, such as N1-methylpseudouridine (m1Ψ) to the mRNA [18,19] (Figure 1). The coding region of the mRNA is flanked by two untranslated regions (UTRs) followed by a polyadenylated (polyA) tail at 3' and a cap at 5' for further structural stabilization and protection (Figure 1) [17,19,20].
As mRNA vaccines are based on pre-fusion epitopes, the fusion pathology may be undeterred, allowing viral infection by syncytia formation to continue unabated [3,16]. This is significant, as it could account for the reoccurrence of COVID-19 symptoms in fully vaccinated individuals [3,21,22]. In addition, this may explain the rare post-vaccination events associated with cell-cell fusion, including giant cell myocarditis, giant cell arthritis, and Creutzfeldt-Jakob Disease, recorded in Vaccine Adverse Event Reporting System (VAERS) database (please see section "Vaccine core: The synthetic mRNA") [23-27].
What is cell-cell fusion?
Cell-cell fusion is a physiological or pathological process in which one or more adjacent cells merge their plasma membranes, cytoplasm, nuclei, and intracellular organelles, generating multinucleated syncytia often with novel, emerging properties [28]. Under normal circumstances, cell-cell fusion occurs during fertilization and placentation as well as during the formation of myoblasts and osteoclasts [29]. Likewise, in the central nervous system (CNS), astrocytes engender physiological syncytia via connexin-mediated cytosolic exchange [30,31].
Several viruses, including SARS-CoV-2, exploit host physiological fusogens as these molecules induce premature cellular senescence and immunosuppressant, phenotypes hospitable to viral replication and, immune evasion [32,33]. Indeed, cell-cell fusion is triggered by ePS, a marker of low immunogenicity, exploited by the SARS-CoV-2 virus to enter into host cells undetected [9,34-36]. Virus-upregulated intracellular calcium (Ca2+) activates transmembrane protein 16F (TMEM16F), that in turn flips PS from the inner to the outer layer of cell membranes, promoting fusion [9].
Viruses, including SARS-CoV-2, can enter host cells via endocytosis or cell-cell fusion, processes driven by cell-penetrating peptides (CPPs), suggesting that endocytosis inhibition may not always prevent infection [37-39]. Endocytosis requires viral protein attachment to a cell surface receptor and internalization of the entire virus/receptor complex. For example, SARS-CoV-2 binds to host angiotensin-converting enzyme-2 (ACE-2) via its S1 protein followed by endocytosis. On the other hand, ePS activates ADAM17, inducing cell-cell fusion via furin cleaving site (FCS) located within the S2 protein, a pathway independent of S1/ACE-2 attachment [40-43].
Vaccine Core: The Synthetic mRNA
In contrast to traditional vaccines that present a plethora of viral proteins to the host immune cells, COVID-19 mRNA-based therapeutics are limited to the antigen of interest (AOI) and elicit antibodies primarily against S1 receptor binding site (RBS) [1,12,44]. For the complete success of the above method, it must be assumed that the SARS-CoV-2 virus cannot ingress host cells by an alternative pathway. However, several studies have highlighted other potential routes of viral ingress, including metalloprotease, integrins, glucose-regulating protein 78 (GRP78), antibody-dependent enhancement, cell-penetrating peptides, and possibly HERV activation [45-47]. These entry points will be discussed in more detail after a brief examination of LNP components.
LNPs
Although transfection data is mostly proprietary, interrogation of LNP components can provide clues about the mRNA ingress into human cells [6]. For example, LNPs contain ionizable lipids, phospholipid 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC), and cholesterol that can attract host phagocytes to internalize the particle [48-50]. Both ionizable lipids and DSPC resemble PS, communicating to phagocytes readiness for engulfment [8]. Aside from comprising an established "eat me" signal, ePS can also convey "fuse me" cues to host phagocytes that can contribute to the unintended consequence of pathological syncytia formation [10]. The LNP component, cholesterol, is also a promoter of pathological cell-cell fusion as it can alter the asymmetry of cell membranes [50]. Moreover, as cell-cell fusion leads to premature cellular senescence and iatrogenic immunosuppression, it may partly explain the immune dysfunction documented in some vaccinated individuals [51-53].
Polyethylene glycol (PEG)
PEG was added to the LNP to stabilize and prolong the mRNA duration of action (Figure 1). The extensive utilization of PEG over the past few decades, suggests that preexisting antibodies could trigger hypersensitivity to vaccines containing this molecule [54,55]. Aside from allergy, PEG is also an established chemical fusogen that can generate pathology by promoting polynucleation, aneuploidy, and genomic instability [56-59]. In addition, PEG up regulates intracellular Ca2+, activating the transmembrane protein 16F (TMEM16F), a lipid scramblase that flips PS on the cell surface, triggering fusion, premature cellular senescence, and immunosuppression [60-63]. As these phenotypes are virus-friendly, PEG-induced cell-cell fusion may inadvertently facilitate not only SARS-CoV-2 but also other viral infections [60,64-66]. Furthermore, ePS-activated ADAM 10 and 17 promote syncytia formation via metalloprotease pathway [40,42,43].
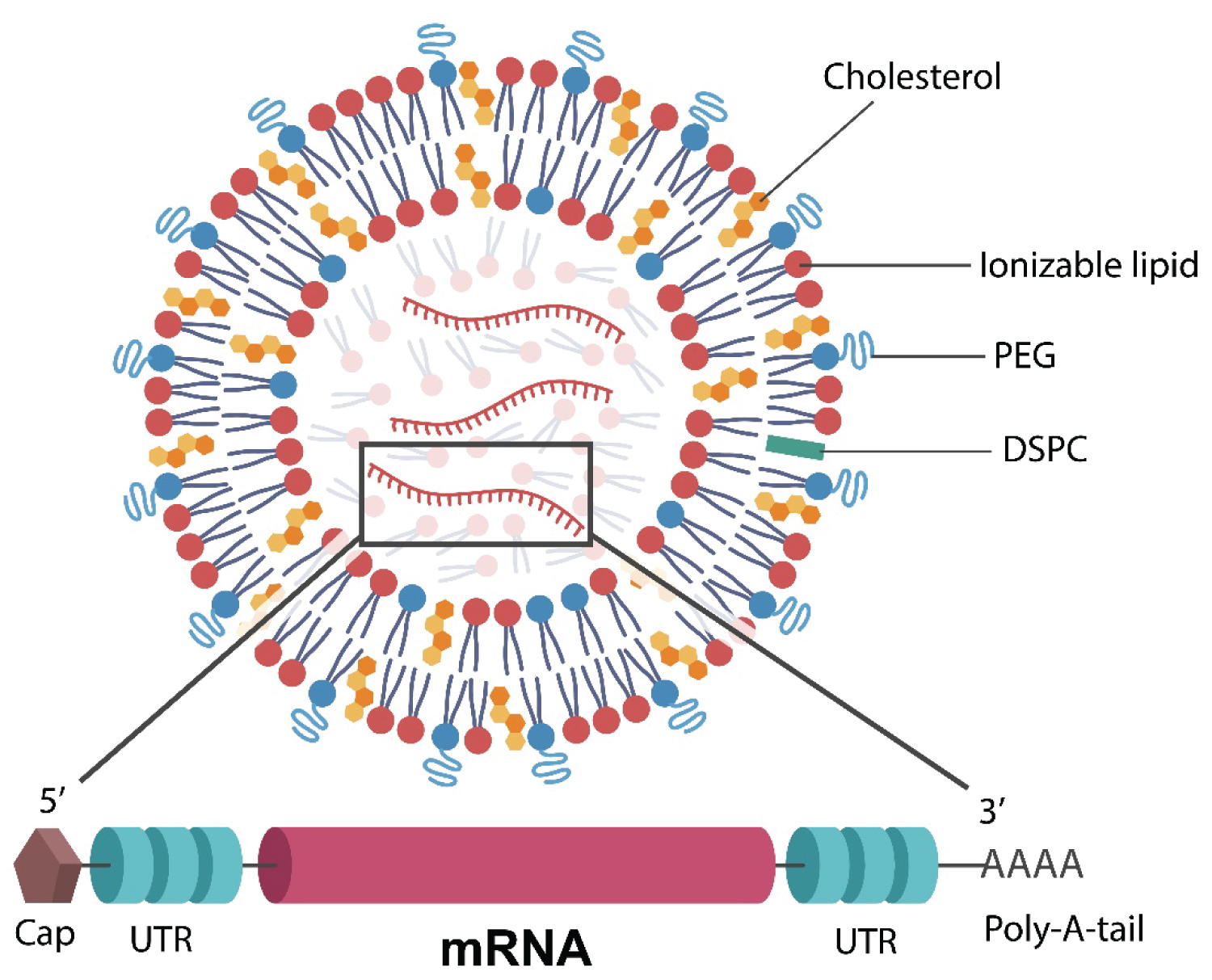 Figure 1: N1-methylpseudouridine (m1Ψ)-modified mRNA (in the rectangle) is surrounded by a lipid nanoparticle (LNP) comprised of 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC), cholesterol, and an ionizable lipid. Polyethylene glycol (PEG) is conjugated with the lipid molecules to increase the mRNA duration of action. The mRNA encodes for the full-length S antigen and is flanked by two untranslated regions (UTRs) and a polyadenylated (polyA) tail at the 3' end for stabilization. A cap at the 5' end offers further protection from exonuclease recognition.
View Figure 1
Figure 1: N1-methylpseudouridine (m1Ψ)-modified mRNA (in the rectangle) is surrounded by a lipid nanoparticle (LNP) comprised of 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC), cholesterol, and an ionizable lipid. Polyethylene glycol (PEG) is conjugated with the lipid molecules to increase the mRNA duration of action. The mRNA encodes for the full-length S antigen and is flanked by two untranslated regions (UTRs) and a polyadenylated (polyA) tail at the 3' end for stabilization. A cap at the 5' end offers further protection from exonuclease recognition.
View Figure 1
PEG was never used in an approved vaccine therefore, its presence in Pfizer-BioNTech and Moderna-1273 therapeutics raised concerns, especially regarding anaphylactic and fusogenic adverse effects [52,67,68]. Moreover, PEG promotes temporary permeabilization of the blood-brain barrier (BBB), a property exploited by the pharmaceutical industry for CNS delivery systems [69-71]. This may account, at least in part, for the rare VAERS-reported neuropsychiatric symptoms, including neurodegenerative disorders [72-74]. Furthermore, earlier studies have demonstrated that PEG may interfere with the conformational stability of proteins, indicating that syncytia, cellular senescence and, dysfunctional proteostasis are highly intertwined [75-77]. While the attention to PEG and the need to further study its relation to the potential vaccine adverse reactions is logical and appropriate, it must be noted that excipients other than PEG might be also be involved in such reported adverse reaction events [78].
DSPC and ionizable lipids
To deliver the liposome cargo to human immune cells, mRNA therapeutics must trick host phagocytes into internalizing LNPs by phagocytosis [79,80]. This is accomplished by decorating the liposomal particles with ionizable lipids and DSPC, an anionic phospholipid that mimics ePS and conveys readiness for phagocytosis [81,82]. This "eat me" signal is exploited by many viruses, including SARS-CoV-2, to enter host immune cells by engulfment [8].
Pfizer and Moderna vaccines were designed to "imitate" dying cells or apoptotic bodies by utilizing ionizable lipids and DSPC delivering mRNA directly to the immune cells' translation machinery [5]. However, DSPC's resemblance to PS may inadvertently activate ADAM 10 and 17, promoting pathological cell-cell fusion and subsequent pathology [40].
Taken together, LNPs mimicking ePS are engulfed by host immune cells, and generate anti-S antibodies by delivering the mRNA cargo to host ribosomes. However, in some cases the disruption of plasma cell asymmetry may inadvertently engender iatrogenic syncytia by activating the metalloprotease pathway.
Potential Non-RBS Modalities of SARS-CoV-2 Infection
In the sections below, we take a closer look at SARS-CoV-2 infection by RBS/ACE-2 independent pathways that may escape neutralization by mRNA vaccines.
Infection by fusion, the metalloprotease pathway
SARS-CoV-2 infection can be disseminated from cell to cell by pathological syncytia, a FCS-dependent route also known as the metalloprotease pathway [83,84]. This infection modality may promote higher infectivity than the RBS/ACE-2 route as FCS deletion was demonstrated to attenuate infectivity [85]. Moreover, FCS activates ADAM 10 and 17, master regulators of cell-cell fusion, enhancing both syncytia formation and contagiousness [11,12]. Thus, the metalloprotease pathway may be resistant to mRNA vaccines that were designed primarily to neutralizepre-fusion epitopes, likely explaining the emergence of COVID-19 symptoms in fully vaccinated individuals [2,3,40,42,43,86]. Furthermore, as pathological cell-cell fusion was associated with premature cellular senescence and immunosuppression, the metalloprotease pathway may account for the dysfunctional immune responses observed in some vaccinated individuals [35,36,51]. Syncytia-related pathology may also contribute to other VAERs-documented post-vaccination events, including giant cell myocarditis, arthritis, and neurodegeneration [23,24,27,87].
Antibody-dependent enhancement
Antibody-dependent enhancement (ADE) is a mechanism of increased viral infection in the presence of partially neutralizing antibodies that can activate immunoglobulin G (IgG) Fc-gamma receptors (FcγRs) [88-91]. This "Trojan horse" infection modality occurs when the SARS-CoV-2 virus hijacks antibodies to infect immune cells and decrease host antiviral defenses [92,93]. This infection route is routinely employed by many viruses, including the human cytomegalovirus (HCMV) known for usurping host T lymphocytes and macrophages [94-97].
The ADE infection pathway presents with the following characteristics: 1. Direct correlation with the disease severity, 2. Independent of S1/ACE-2 attachment, and 3. Probably unaffected by the mRNA vaccines [94,98].
Cell-penetrating peptides
COVID-19 proteomic studies show that SARS-CoV-2 expresses many cell-penetrating peptides (CPP) that can promote viral entry and may be undeterred by the mRNA vaccines [99]. Indeed, many enveloped and unenveloped viruses contain CPPs and employ them for ingress host cells [100].
Aside from SARS-CoV-2, several other viruses, including the H5N1 avian influenza, dengue virus, and human papilloma virus can enter host cells via CPPs, indicating a common viral entry route [101,102]. In addition, CPPs are being utilized as pharmacological vehicles for intracellular delivery of therapeutics, highlighting the capability of these molecules to cross cell membranes [103]. CPPs can up regulate intracellular Ca2+, promoting both pathological syncytia and protein misfolding [26,104-106].
Viroporins
Viroporins are hydrophobic, voltage-independent viral proteins known for piercing plasma membranes, triggering cell death. Many viruses, including SARS-CoV-2, express viroporins and promote infectivity, as these proteins are known for mediating viral entry and exit [107,108]. A recent in silico study found that the SARS-CoV-2 virus expresses three viroporins, the E antigen, open reading frame 3a (ORF3a), and ORF8a, highlighting potential, non-RBS, routes of viral ingress that may be refractory to mRNA vaccines [109]. Interestingly, vaccine-mediated neutralization of S1 protein may contribute to the accumulation of other viral proteins, including viroporins, in the extracellular space (ECS), probably opening alternative entry portals for viral ingress [108,110]. In addition, viroporin channels disrupt the ionic homeostasis of host cells, up regulating intracellular Ca2+ that in turn, promotes pathological cell-cell fusion [111].
HERVs
FCS was reported to activate HERVs, primarily type W and K, triggering not only cell-cell fusion but also hyperinflammatory responses and dysfunctional proteostasis [112-115]. HERVs are viral fossils embedded in human DNA that can be "awakened" by the infection with exogenous viruses, cancer, or neuropsychiatric conditions [116-118].
Several studies demonstrated that SARS-CoV-2 can activate HERV-W, an ancestral gene that encodes for the physiological placental fusogen syncytin-1 responsible for the merger of trophoblasts during the early pregnancy [112,113]. This suggests that the reproductive post-vaccine events may be triggered by the FCS pathology. Virus-up regulated syncytin-1 may promote aberrant cell-cell fusion throughout the host tissues and organs, including the brain [119,120]. Interestingly, Omicron variant convalescent sera contain anti-FCS antibodies, suggesting that it may neutralize not only the pathological cell-cell fusion but also HERV activation [21,121].
Conclusions
The mRNA vaccines were approved on an emergency basis to combat COVID-19 pandemic. These vaccines also represent the first administration of LNPs at large scale. Taken together, they constitute milestones in the development of a novel and much promising therapeutics delivery field. Having said that, at the time of the emergency approval, the S2 viral antigen was insufficiently studied, and the FCS-mediated fusion pathology was mostly unknown. These aspects are now starting to attract attention, in an effort to best understand the underlying cellular mechanisms, pathways and potential unintended consequences.
The Pfizer and Moderna vaccines elicit powerful neutralizing antibodies against the RBS located on S1 protein and block viral entry by endocytosis. However, the S2-dependent metalloprotease pathway and other potential entry portals may not be adequately addressed by these therapeutics. Residual COVID-19 symptoms, often conceptualized as vaccine adverse effects, could be caused by FCS-mediated pathology. ADAM inhibitors and/or Omicron convalescent sera may effectively eradicate the SARS-CoV-2 virus by inhibiting metalloprotease pathway.
Disclaimer
Where authors are identified as personnel of the International Agency for Research on Cancer/WHO, the authors alone are responsible for the views expressed in this article and they do not necessarily represent the decisions, policy or views of the International Agency for Research on Cancer/WHO.
References
-
Rijkers GT, Weterings N, Obregon-Henao A, Lepolder M, Dutt TS, et al. (2021) Antigen presentation of mRNA-based and virus-vectored SARS-CoV-2 vaccines. Vaccines (Basel) 9: 848.
-
Bergwerk M, Gonen T, Lustig Y, Amit S, Lipsitch M, et al. (2021) Covid-19 breakthrough infections in vaccinated health care workers. N Engl J Med 385: 1474-1484.
-
Hein S, Benz NI, Eisert J, Herrlein ML, Oberle D, et al. (2021) Comirnaty-elicited and convalescent sera recognize different spike epitopes. Vaccines (Basel) 9: 1419.
-
Sakai-Kato K, Yoshida K, Takechi-Haraya Y, Izutsu KI (2020) Physicochemical characterization of liposomes that mimic the lipid composition of exosomes for effective intracellular trafficking. Langmuir 36: 12735-12744.
-
Lotter C, Alter CL, Bolten JS, Detampel P, Palivan CG, et al. (2022) Incorporation of phosphatidylserine improves efficiency of lipid based gene delivery systems. Eur J Pharm Biopharm 172: 134-143.
-
Cao Y, Gao GF (2021) mRNA vaccines: A matter of delivery. Clinical Medicine 32: 100746.
-
Liu J, Conboy JC (2009) Phase behavior of planar supported lipid membranes composed of cholesterol and 1,2-Distearoyl-sn-Glycerol-3-Phosphocholine examined by sum-frequency vibrational spectroscopy. VibSpectrosc 50: 106-115.
-
Bohan D, Ert HV, Ruggio N, Rogers KJ, Badreddine M, et al. (2021) Phosphatidylserine receptors enhance SARS-CoV-2 infection: AXL as a therapeutic target for COVID-19. bioRxiv.
-
Whitlock JM, Chernomordik LV (2021) Flagging fusion: Phosphatidylserine signaling in cell-cell fusion. J Biol Chem 296: 100411.
-
Kim GW, Park SY, Kim IS (2016) Novel function of stabilin-2 in myoblast fusion: The recognition of extracellular phosphatidylserine as a "fuse-me" signal. BMB Rep 49: 303-304.
-
Yamamoto M, Gohda J, Kobayashi A, Tomita K, Hirayama Y, et al. (2022) Metalloproteinase-dependent and TMPRSS2-independent cell surface entry pathway of SARS-CoV-2 requires the furin cleavage site and the S2 domain of spike protein. mBio 13: e0051922.
-
Theuerkauf SA, Michels A, Riechert V, Maier TJ, Flory E, et al. (2021) Quantitative assays reveal cell fusion at minimal levels of SARS-CoV-2 spike protein and fusion from without. iScience 24: 102170.
-
Pardi N, Hogan MJ, Porter FW, Weissman D (2018) mRNA vaccines - a new era in vaccinology. Nat Rev Drug Discov 17: 261-279.
-
Gokita K, Inoue J, Ishihara H, Kojima K, Inazawa J (2020) Therapeutic potential of LNP-mediated delivery of miR-634 for cancer therapy. Mol Ther Nucleic Acids 2020 19: 330-338.
-
Kulkarni JA, Cullis PR, van der Meel R (2018) Lipid nanoparticles enabling gene therapies: From concepts to clinical utility. Nucleic Acid Ther 28: 146-157.
-
Andries O, Mc Cafferty S, De Smedt SC, Weiss R, Sanders NN, et al. (2015) N(1)-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. J Control Release 217: 337-344.
-
Schoenmaker L, Witzigmann D, Kulkarni JA, Verbeke R, Kersten G, et al. (2021) mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int J Pharm 601: 120586.
-
Nance KD, Meier JL (2021) Modifications in an Emergency: The role of N1-Methylpseudouridine in COVID-19 vaccines. ACS Cent Sci 7: 748-756.
-
Gebre MS, Rauch S, Roth N, Yu J, Chandrashekar A, et al. (2022) Optimization of non-coding regions for a non-modified mRNA COVID-19 vaccine. Nature 601: 410-414.
-
Jeeva S, Kim KH, Shin CH, Wang BZ, Kang SM (2021) An update on mRNA-based viral vaccines. Vaccines (Basel) 9: 965.
-
Hu J, Chen X, Lu X, Wu L, Yin L, et al. (2022) A spike protein S2 antibody efficiently neutralizes the Omicron variant. Cell Mol Immunol 19: 644-646.
-
Ren F, Zhang N, Zhang L, Miller E, Pu JJ (2020) Alternative polyadenylation: A new frontier in post transcriptional regulation. Biomark Res 8: 67.
-
Sung K, McCain J, King KR, Hong K, Aisagbonhi O, et al. (2022) Biopsy-proven giant cell myocarditis following the COVID-19 vaccine. Circ Heart Fail 15: e009321.
-
Lazebnik Y (2021) Cell fusion as a link between the SARS-CoV-2 spike protein, COVID-19 complications, and vaccine side effects. Oncotarget 12: 2476-2488.
-
Anzola AM, Trives L, Martínez-Barrio J, Pinilla B, Álvaro-Gracia JM, et al. (2022) New-onset giant cell arteritis following COVID-19 mRNA (BioNTech/Pfizer) vaccine: A double-edged sword? Clin Rheumatol 41: 1623-1625.
-
Kidson C, Moreau MC, Asher DM, Brown PW, Coon HG, et al. (1978) Cell fusion induced by scrapie and Creutzfeldt-Jakob virus-infected brain preparations. Proc Natl Acad Sci USA 75: 2969-2971.
-
Perez JC, Moret-Chalmin C, Montagnier L (2022) Towards the emergence of a new form of the neurodegenerative Creutzfeldt-Jakob disease: Twenty six cases of CJD declared a few days after a COVID-19 “vaccine” Jab.
-
Lin L, Li Q, Wang Y, Shi Y (2021) Syncytia formation during SARS-CoV-2 lung infection: A disastrous unity to eliminate lymphocytes. Cell Death Differ 28: 2019-2021.
-
Kar M, Ghosh D, Sengupta J (2007) Molecular correlates of syncytialization in muscle and placenta. Indian J Physiol Pharmacol 51: 311-325.
-
Refaeli R, Doron A, Benmelech-Chovav A, Groysman M, Kreisel T, et al. (2021) Features of hippocampal astrocytic domains and their spatial relation to excitatory and inhibitory neurons. Glia 69: 2378-2390.
-
Katoozi S, Skauli N, Zahl S, Deshpande T, Ezan P, et al. (2020) Uncoupling of the astrocyte syncytium differentially affects AQP4 isoforms. Cells 9: 382.
-
de Souza Cardoso R, Viana RMM, Vitti BC, Coelho ACL, de Jesus BLS, et al. (2021) Human respiratory syncytial virus infection in a human T cell line is hampered at multiple steps. Viruses 13: 231.
-
Vance TDR, Lee JE (2020) Virus and eukaryote fusogen superfamilies. Curr Biol 30: R750-R754.
-
Hörnich BF, Großkopf AK, Schlagowski S, Tenbusch M, Kleine-Weber H, et al. (2021) SARS-CoV-2 and SARS-CoV spike-mediated cell-cell fusion differ in their requirements for receptor expression and proteolytic activation. J Virol 95: e00002-e00021.
-
Birge R, Boeltz S, Kumar S, Carlson J, Wanderley J, et al. (2016) Phosphatidylserine is a global immunosuppressive signal in efferocytosis, infectious disease, and cancer. Cell Death Differ 23: 962-978.
-
Gal H, Krizhanovsky V (2014) Cell fusion induced senescence. Aging (Albany NY) 6: 353-354.
-
Chou T (2007) Stochastic entry of enveloped viruses: Fusion versus endocytosis. Biophys J 93: 1116-1123.
-
Kumar CS, Dey D, Ghosh S, Banerjee M (2018) Breach: Host membrane penetration and entry by nonenveloped viruses. Trends Microbiol 26: 525-537.
-
Yamauchi Y, Greber UF (2016) Principles of virus uncoating: Cues and the snooker ball. Traffic 17: 569-592.
-
Sommer A, Kordowski F, Büch J, Maretzky T, Evers A, et al. (2016) Phosphatidylserine exposure is required for ADAM17 sheddase function. Nat Commun 7: 11523.
-
Bartolomé A, Liang J, Wang P, Ho DD, Pajvani UP, et al. (2021) Angiotensin converting enzyme 2 is a novel target of the γ-secretase complex. Sci Rep 11: 9803.
-
Jocher G, Grass V, Tschirner SK, Riepler L, Breimann S, et al. (2022) ADAM10 and ADAM17 promote SARS-CoV-2 cell entry and spike protein-mediated lung cell fusion. EMBO Rep 23: e54305.
-
de Seabra Rodrigues Dias IR, Cao Z, Kwok HF (2022) Adamalysins in COVID-19 - Potential mechanisms behind exacerbating the disease. Biomed Pharmacother 150: 112970.
-
Zeng C, Evans JP, King T, Zheng Yi-Min, Oltz EM, et al. (2021) SARS-CoV-2 spreads through cell-to-cell transmission. PNAS 119: e2111400119.
-
Allam L, Ghrifi F, Mohammed H, El Hafidi N, El Jaoudi R, et al. (2020) Targeting the GRP78-dependant SARS-CoV-2 cell entry by peptides and small molecules. Bioinform Biol Insights 14: 1177932220965505.
-
Puray-Chavez M, LaPak KM, Schrank TP, Elliott JL, Bhatt DP, et al. (2021) Systematic analysis of SARS-CoV-2 infection of an ACE2-negative human airway cell. Cell Rep 36: 109364.
-
Sigrist CJ, Bridge A, Le Mercier P (2020) A potential role for integrins in host cell entry by SARS-CoV-2. Antiviral Res 177: 104759.
-
Horejs C (2021) From lipids to lipid nanoparticles to mRNA vaccines. Nat Rev Mater 6: 1075-1076.
-
Maugeri M, Nawaz M, Papadimitriou A, Angerfors A, Camponeschi A, et al. (2019) Linkage between endosomal escape of LNP-mRNA and loading into EVs for transport to other cells. Nat Commun 10: 4333.
-
Yang ST, Kreutzberger AJB, Lee J, Kiessling V, Tamm LK (2016) The role of cholesterol in membrane fusion. Chem Phys Lipids 199: 136-143.
-
Seneff S, Nigh G, Kyriakopoulos AM, McCullough PA (2022) Innate immune suppression by SARS-CoV-2 mRNA vaccinations: The role of G-quadruplexes, exosomes, and MicroRNAs. Food Chem Toxicol 164: 113008.
-
Yang J, Shen MH (2006) Polyethylene glycol-mediated cell fusion. Methods Mol Biol 325: 59-66.
-
Kessler E, Kambhampati L, Kill G, Mackenzie E, Meyer M, et al. (2020) The prion-like cytoplasmic polyadenylation element binding (CPEB) protein: Can protein aggregations in long-term memory formation inform memory loss in amyloid associated neurodegenerative disease? The FASEB Journal 34.
-
de Vrieze J (2020) Suspicions grow that nanoparticles in Pfizer’s COVID-19 vaccine trigger rare allergic reactions. Science.
-
Alconcel SNS, Baas AS, Maynard HD (2011) FDA-approved poly (ethylene glycol)-protein conjugate drugs. Polym Chem 2: 1442-1448.
-
Chen SL, Fu RH, Liao SF, Liu SP, Lin SZ, et al. (2018) A PEG-based hydrogel for effective wound care management. Cell Transplant 27: 275-284.
-
Kim CY, Sikkema WK, Hwang IK, Oh H, Kim UJ, et al. (2016) Spinal cord fusion with PEG-GNRs (TexasPEG): Neurophysiological recovery in 24 hours in rats. Surg Neurol Int 7: S632-S636.
-
Lane RD, Crissman RS, Lachman MF (1984) Comparison of polyethylene glycols as fusogens for producing lymphocyte-myeloma hybrids. J Immunol Methods 72: 71-76.
-
Sfera A, Sasannia S, Kozlakidis Z (2022) PEGylated COVID-19 vaccines and cell-cell fusion. Arch Clin Trials 2: 1-2.
-
Tordoff MG (1997) Polyethylene glycol-induced calcium appetite. Am J Physiol 1997 273: R587-R596.
-
Kucherenko Iu V (1999) Intracellular free calcium content increase in the erythrocytes treated with the cryoprotective medium based on polyethylene glycol 1500 (PEG-1500)]. Ukr Biokhim Zh 80: 124-130.
-
Wu N, Cernysiov V, Davidson D, Song H, Tang J, et al. (2020) Critical role of lipid scramblase TMEM16F in phosphatidylserine exposure and repair of plasma membrane after pore formation. Cell Rep 30: 1129-1140.
-
Pannuzzo M, De Jong DH, Raudino A, Marrink SJ (2014) Simulation of polyethylene glycol and calcium-mediated membrane fusion. J Chem Phys 140: 124905.
-
Arudchandran R, Brown MJ, Song JS, Wank SA, Haleem-Smith H, et al. (1999) Polyethylene glycol-mediated infection of non-permissive mammalian cells with semliki forest virus: Application to signal transduction studies. J Immunol Methods 222: 197-208.
-
Gripon P, Diot C, Guguen-Guillouzo C (1993) Reproducible high level infection of cultured adult human hepatocytes by hepatitis B virus: Effect of polyethylene glycol on adsorption and penetration. Virology 192: 534-540.
-
Walker EB, Pritchard SM, Cunha CW, Aguilar HC, Nicola AV (2015) Polyethylene glycol-mediated fusion of herpes simplex type 1 virions with the plasma membrane of cells that support endocytic entry. Virol J 12: 190.
-
Lentz BR (2007) PEG as a tool to gain insight into membrane fusion. Eur Biophys J 36: 315-326.
-
Zhou ZH, Stone CA Jr, Jakubovic B, Phillips EJ, Sussman G, et al. (2020) Anti-PEG IgE in anaphylaxis associated with polyethylene glycol. J Allergy Clin Immunol Pract 9: 1731-1733.
-
Rabanel JM, Piec PA, Landri S, Patten SA, Ramassamy C (2020) Transport of PEGylated-PLA nanoparticles across a blood brain barrier model, entry into neuronal cells and in vivo brain bioavailability. J Control Release 328: 679-695.
-
Fernandes C, Pinto M, Martins C, Gomes MJ, Sarmento B, et al. (2018) Development of a PEGylated-based platform for efficient delivery of dietary antioxidants across the blood-brain barrier. Bioconjug Chem 29: 1677-1689.
-
Paskal AM, Paskal W, Pietruski P, Wlodarski PK (2019) Polyethylene glycol: The future of posttraumatic nerve Repair? systemic review. Int J Mol Sci 20: 1478.
-
Einstein EH, Shahzadi A, Desir L, Katz J, Boockvar J, et al. (2021) New-onset neurologic symptoms and related neuro-oncologic lesions discovered after COVID-19 vaccination: Two neurosurgical cases and review of post-vaccine inflammatory responses. Cureus 13: e15664.
-
Frontera JA, Tamborska AA, Doheim MF, Garcia-Azorin D, Gezegen H, et al. (2022) Neurological events reported after COVID-19 vaccines: An analysis of VAERS. Ann Neurol 91: 756-771.
-
Assiri SA, Althaqafi RMM, Alswat K, Alghamdi AA, Alomairi NE, et al. (2022) Post COVID-19 vaccination-associated neurological complications. Neuropsychiatr Dis Treat 18: 137-154.
-
Wälchli R, Fanizzi F, Massant J, Arosio P (2020) Relationship of PEG-induced precipitation with protein-protein interactions and aggregation rates of high concentration mAb formulations at 5 °C. Eur J Pharm Biopharm 151: 53-60.
-
Meng W, Guo X, Qin M, Pan H, Cao Y, et al. (2012) Mechanistic insights into the stabilization of srcSH3 by PEGylation. Langmuir 28: 16133-16140.
-
Monfardini C, Schiavon O, Caliceti P, Morpurgo M, Harris JM, et al. (1995) A branched monomethoxypoly (ethylene glycol) for protein modification. Bioconjugate Chem 6: 62-69.
-
Borgsteede SD, Geersing TH, Tempels-Pavlica Ž (2021) Other excipients than PEG might cause serious hypersensitivity reactions in COVID-19 vaccines. Allergy 76: 1941-1942.
-
Ter Braake D, Benne N, Lau CYJ, Mastrobattista E, Broere F (2021) Retinoic acid-containing liposomes for the induction of antigen-specific regulatory T cells as a treatment for autoimmune diseases. Pharmaceutics 13: 1949.
-
Nijen T wilhaar MK, Czentner L, van Nostrum CF, Storm G, den Haan JMM (2021) Mimicking pathogens to augment the potency of liposomal cancer vaccines. Pharmaceutics 13: 954.
-
Segawa K, Nagata S (2015) An apoptotic 'Eat Me' signal: Phosphatidylserine exposure. Trends Cell Biol 25: 639-650.
-
Norling K, Bernasconi V, Agmo Hernández V, Parveen N, Edwards K, et al. (2019) Gel phase 1,2-Distearoyl-sn-glycero-3-phosphocholine-based liposomes are superior to fluid phase liposomes at augmenting both antigen presentation on major histocompatibility complex class ii and costimulatory molecule display by dendritic cells in vitro. ACS Infect Dis 5: 1867-1878.
-
Johnson BA, Xie X, Kalveram B, Lokugamage KG, Muruato A, et al. (2020) Furin cleavage site is key to SARS-CoV-2 pathogenesis. bioRxiv.
-
Peacock TP, Goldhill DH, Zhou J, Baillon L, Frise R, et al. (2021) The furin cleavage site in the SARS-CoV-2 spike protein is required for transmission in ferrets. Nat Microbiol 6: 899-909.
-
Wang P, Lau SY, Deng S, Chen P, Yee Mok BW, et al. (2021) Characterization of an attenuated SARS-CoV-2 variant with a deletion at the S1/S2 junction of the spike protein. Nat Commun 12.
-
Abu-Raddad LJ, Chemaitelly H, Ayoub HH, Tang P, Coyle P, et al. (2022) Relative infectiousness of SARS-CoV-2 vaccine breakthrough infections, reinfections, and primary infections. Nat Commun 13: 532.
-
Trougakos IP, Terpos E, Alexopoulos H, Politou M, Paraskevis D, et al. (2022) Adverse effects of COVID-19 mRNA vaccines: the spike hypothesis. Trends Mol Med 28: 542-554.
-
Bournazos S, Gupta A, Ravetch JV (2020) The role of IgG Fc receptors in antibody-dependent enhancement. Nat Rev Immunol 20: 633-643.
-
Arvin AM, Fink K, Schmid MA, Cathcart A, Spreafico R, et al. (2020) A perspective on potential antibody-dependent enhancement of SARS-CoV-2. Nature 584: 353-363.
-
Wieczorek Lindsay, Zemil M, Merbah M, Dussupt V, Kavusak E, et al. (2022) Evaluation of antibody-dependent fc-mediated viral entry, as compared with neutralization, in SARS-CoV-2 infection. Front Immunol 13.
-
Wan Y, Shang J, Sun S, Tai w, Chen J, et al. (2020) Molecular mechanism for antibody-dependent enhancement of coronavirus entry. J Virol 94: e02015-e02019.
-
Maemura T, Kuroda M, Armbrust T, Yamayoshi S, Halfmann PJ, et al. (2021) Antibody-dependent enhancement of SARS-CoV-2 infection is mediated by the IgG receptors FcγRIIA and FcγRIIIA but does not contribute to aberrant cytokine production by macrophages. mBio 12: e0198721.
-
Kulkarni R (2020) Antibody-dependent enhancement of viral infections. In Dynamics of Immune Activation in Viral Diseases, 9-41.
-
Shen XR, Geng R, Li Q, Chen Y, Li SF, et al. (2022) ACE2-independent infection of T lymphocytes by SARS-CoV-2. Sig Transduct Target Ther 7.
-
Kolb P, Hoffmann K, Sievert A, Reinhard H, Merce-Maldonado E, et al. (2021) Human cytomegalovirus antagonizes activation of Fcγ receptors by distinct and synergizing modes of IgG manipulation. Elife 10: e63877.
-
Farouq MAH, Acevedo R, Ferro VA, Mulheran PA, Al Qaraghuli MM (2022) The role of antibodies in the treatment of SARS-CoV-2 virus infection, and evaluating their contribution to antibody-dependent enhancement of infection. Int J Mol Sci 23: 6078.
-
Sefik E, Qu R, Junqueira C, Kaffe E, Mirza H, et al. (2022) Inflammasome activation in infected macrophages drives COVID-19 pathology. Nature 606: 585-593.
-
Partridge LJ, Urwin L, Nicklin MJH, James DC, Green LR, et al. (2021) ACE2-independent interaction of SARS-CoV-2 spike protein with human epithelial cells is inhibited by unfractionated heparin. Cells 10: 1419.
-
Hemmati S, Behzadipour Y, Haddad M (2020) Decoding the proteome of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) for cell-penetrating peptides involved in pathogenesis or applicable as drug delivery vectors. Infect Genet Evol 85.
-
Qi X, Droste T, Kao CC (2011) Cell-penetrating peptides derived from viral capsid proteins. Mol Plant Microbe Interact 24: 25-36.
-
Freire JM, Veiga AS, Rego de Figueiredo I, de la Torre BG, Santos NC, et al. (2014) Nucleic acid delivery by cell penetrating peptides derived from dengue virus capsid protein: Design and mechanism of action. FEBS J 281: 191-215.
-
Kajiwara N, Nomura N, Ukaji M, Yamamoto N, Kohara M, et al. (2020) Cell-penetrating peptide-mediated cell entry of H5N1 highly pathogenic avian influenza virus. Sci Rep 10.
-
Zhang P, Monteiro da Silva G, Deatherage C, Burd C, DiMaio D (2018) Cell-penetrating peptide mediates intracellular membrane passage of human papillomavirus L2 protein to trigger retrograde trafficking. Cell 174: 1465-1476.
-
Liu S, Hossinger A, Heumüller SE, Hornberger A, Buravlova O, et al. (2021) Highly efficient intercellular spreading of protein misfolding mediated by viral ligand-receptor interactions. Nat Commun 12.
-
Aoe T (2020) Pathological aspects of COVID-19 as a conformational disease and the use of pharmacological chaperones as a potential therapeutic strategy. Front Pharmacol 11.
-
Gessner I, Neundorf I (2020) Nanoparticles modified with cell-penetrating peptides: Conjugation mechanisms, physicochemical properties, and application in cancer diagnosis and therapy. Int J Mol Sci 21: 2536.
-
Nieva J, Madan V, Carrasco L (2012) Viroporins: Structure and biological functions. Nat Rev Microbiol 10: 563-574.
-
Giorda KM, Hebert DN (2013) Viroporins customize host cells for efficient viral propagation. DNA Cell Biol 32: 557-564.
-
Rouka E, Gourgoulianis KI, Zarogiannis SG (2021) In silico investigation of the viroporin E as a vaccine target against SARS-CoV-2. Am J Physiol Lung Cell Mol Physiol 320: L1057-L1063.
-
Nieto-Torres JL, Verdiá-Báguena C, Castaño-Rodriguez C, Aguilella VM, Enjuanes L (2015) Relevance of viroporin ion channel activity on viral replication and pathogenesis. Viruses 7: 3552-3573.
-
Hyser JM, Estes MK (2015) Pathophysiological consequences of calcium-conducting viroporins. Annu Rev Virol 2: 473-496.
-
Durnaoglu S, Lee SK, Ahnn J (2022) Syncytin, envelope protein of human endogenous retrovirus (HERV): No longer 'fossil' in human genome. Anim Cells Syst (Seoul) 25: 358-368.
-
Charvet B, Brunel J, Pierquin J, Iampietro M, Decimo D, et al. (2022) SARS-CoV-2 induces human endogenous retrovirus type W envelope protein expression in blood lymphocytes and in tissues of COVID-19 patients. medRxiv.
-
Grandi N, Tramontano E (2017) Type W human endogenous retrovirus (HERV-W) integrations and their mobilization by L1 machinery: Contribution to the human transcriptome and impact on the host physiopathology. Viruses 9: 162.
-
Lipton SA, Gu Z, Nakamura T (2007) Inflammatory mediators leading to protein misfolding and uncompetitive/fast off-rate drug therapy for neurodegenerative disorders. Int Rev Neurobiol 82: 1-27.
-
Slokar G, Hasler G (2016) Human endogenous retroviruses as pathogenic factors in the development of schizophrenia. Front Psychiatry 6: 183.
-
Dittmar T, Weiler J, Luo T, Hass R (2021) Cell-cell fusion mediated by viruses and HERV-derived fusogens in cancer initiation and progression. Cancers (Basel) 13: 5363.
-
Dolei A, Ibba G, Piu C, Serra C (2019) Expression of HERV Genes as possible biomarker and target in neurodegenerative diseases. Int J Mol Sci 20: 3706.
-
Wang X, Huang J, Zhu F (2018) Human endogenous retroviral envelope protein syncytin-1 and inflammatory abnormalities in neuropsychological diseases. Front Psychiatry 9: 422.
-
Giordano-Santini R, Linton C, Hilliard MA (2016) Cell-cell fusion in the nervous system: Alternative mechanisms of development, injury, and repair. Semin Cell Dev Biol 60: 146-154.
-
Yao L, Zhu KL, Jiang XL, Wang XJ, Zhan BD, et al. (2022) Omicron subvariants escape antibodies elicited by vaccination and BA.2.2 infection. Lancet Infect Dis 22: 1116-1117.
Citation
Sfera A, Thomas KG, Sfera DO, Anton JJ, Andronescu CV, et al. (2022) Do Messenger RNA Vaccines Induce Pathological Syncytia?. Int J Pathol Clin Res 8:137. doi.org/10.23937/2469-5807/1510137