Chlorpromazine hydrochloride (CPZ) refers to antipsychotic drugs. CPZ is drug of first choice for the treatment of schizophrenia. However, its impact on the main excitatory neurotransmitter system, glutamatergic, is unclear. This issue is the aim of our study. Experiments carried out on brain slices of the olfactory cortex of rats. We used electrophysiological techniques extracellular recordings in brain slice to investigate CPZ effects on the activity of the glutamatergic ionotropic alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic receptors (AMPAR) and N-methyl-D-aspartic acid type glutamate receptors (NMDAR). Application CPZ at different concentrations modified activities the both AMPAR and NMDAR. These reactions were phase: Initial activation and subsequent depression for AMPAR at concentrations of CPZ 10-6 M and 10-4 M. For the NMDAR at low concentrations, CPZ (10-6 M and 10-5 M) was the initial activation, and at the end of exposure, these receptors were depressed. In the test of reversibility at washing the activation AMPAR remained elevated and was significantly different from control values. Activity NMDAR returned to the control values at washing. In order to improve functions of NMDAR slices pretreated by ammonium chloride (NH4Cl, 20 mM) which is commonly recognized as fast and efficient lysosomal inhibitor. Such processing of slices and subsequent action of CPZ protected the functioning of NMDAR. Thus, data presented in this study on the influence of CPZ on the ionotropic glutamatergic mechanisms allow understanding the multifaceted mechanisms action of CPZ in protection the deterioration of mental disorders such as schizophrenia.
Chlorpromazine, Schizophrenia, Brain slices, Field potential, Ammonium chloride
Chlorpromazine (CPZ) is on the World Health Organization list of essential medicines remains drug of first choice for the treatment of schizophrenia despite the emergence of other drugs [1,2].
Chlorpromazine hydrochloride (some synonyms aminazine, largactil, thorazine, etc.), the most commonly used form of CPZ in the clinic, refers to a group of antipsychotic drugs. It is a dimethylamine derivative of phenothiazine, has a chemical formula of C17H20Cl2N2S and molecular weight: 355.321 g/mol. In the blood of warm-blooded, it interacts well with plasma proteins, its protein binding reaches 90-99%. The time of its half-life was 30 ± 7 hours, which indicates its long-term stability [3].
The principal pharmacological actions are antipsychotropic. It also exerts sedative and antiemetic activity. CPZ has actions at all levels of the central nervous system, primarily at subcortical structures of the brain - reticular formation of the midbrain. CPZ has strong antiadrenergic and weak peripheral anticholinergic activity. It also possesses slight antihistaminic and antiserotoninergic activity [4,5].
Currently interaction CPZ with dopamine receptors is the dominant view. At the same time its impact on the main excitatory neurotransmitter system, glutamatergic, has hardly been studied, although some fragmentary data confirm such interactions [6]. The importance of this question consists in that in recent years developed intensively theory pathogenesis of schizophrenia, where the main role in this disease play a disturbances glutamatergic mediator system. Confirmation "glutamatergic" hypothesis is based on the fact that the introduction animal NMDA-dependent antagonists of glutamate receptors (phencyclidine, ketamine, MK-801, etc.) is one of the most common and adequate model of schizophrenia [7]. NMDA-receptor antagonists induce behavioral disorders in humans, which are very similar to the clinical manifestations of schizophrenia [8]. Phencyclidine injections can cause memory impairment without evidence of psychosis. Chronic exposure NMDA-antagonists leads to cognitive deficits (as in schizophrenia) [9], which are believed to be associated with dysfunction of NMDAR [10].
Insufficient the specific data on mechanisms of influence of CPZ on glutamatergic receptor mechanisms prompted us to investigate CPZ effects on the activity of key glutamatergic ionotropic receptors, the alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic (AMPAR) and N-methyl-D-aspartic acid type glutamate receptors (NMDAR) [11] of these, AMPAR implement rapid transmission synaptic excitation throughout most of the vertebrate central nervous system. The NMDAR family has received special attention because of its distinct role in the regulation of synaptic plasticity - long-term potentiation/depression and experience-dependent synaptic refinement [12] and because of its critical role in neurological and psychiatric disorders [13]. Hypo- or hyper-activation of NMDAR is critically involved in pain amplification, stroke, epilepsy, schizophrenia, post-traumatic stress disorder, dementia, depression and various neurodegenerative diseases (e.g. Alzheimer's and Parkinson's) [13].
For the experimental study, the influence of CPZ on AMPAR and NMDAR is important to choose an adequate and optimal experimental object, in which was not significant influence of the dopaminergic and serotonergic neurotransmitter systems, because CPZ blocks these receptors. Because our studies conducted on slices the olfactory cortex of Wistar rats. The advantages of these slices is that in them are present and actively operate AMPAR and NMDAR as in the base functions of excitation transfer, as well and in more complex processes of learning. Presence of dopaminergic and serotoninergic receptors in these slices is a minimum [14]. These, data allow us to ignore the involvement of these neurotransmitter systems under the action of CPZ and to submit involvement of AMPAR and NMDAR in slices in "pure form".
In addition, in order to understand the initial processes of the CPZ we studied the changes of activity of the both AMPAR and NMDAR in neuronal network at stimulation the lateral olfactory tract, which is the main afferent input to the cells of the olfactory cortex.
Wistar rats with body weight 100-150 g were obtained from vivarium (Pavlov Institute of Physiology, RAS Saint Petersburg, Russia). All experiments were performed in compliance with ethical standards of the Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes.
Studies were performed on male Wistar rats with body weight 100-150 g (vivarium of the Pavlov Institute of Physiology, RAS). In this work were used methods of the slices preparation and their incubation as described in our previous publications [15]. Tangential slices of olfactory cortex 450-500 µm thick were cut from the brain of male rats of Wistar line with body weight 100-150 g.
The prepared slices were transferred to a glass vial and every slice was preincubated for 1 h in 1 mL of artificial Cerebrospinal Fluid (aCSF) at 37 ℃, pH 7.21-7.24. The composition of aCSF was as follows (mM): NaCl - 124.0, KCl - 5.0, CaCl2 - 2.6, KH2PO4 - 1.24, MgSO4 - 1.2, NaHCO3 - 3.0, tris-HCL (pH 7.4) - 23.0, glucose - 10.0. aCSF was equilibrated with O2.
Extracellular Field Potentials (FPs) were evoked using platinum custom-made bipolar stimulating electrodes positioned onto Lateral Olfactory Tract (LOT). Stimulation was applied as the rectangular pulses (duration - 0.1 msec, intensity - 1.2-1.5 V, frequency - 0.003 Hz) using the stimulator ESU-1 (Russia).
The FPs was recorded using a glass microelectrode filled with 1 M NaCl with tip resistance 1-5 MΩ. Signals were registered with an NTO-2 amplifier (Russia), digitized by analog-to-digital converter MD-32 (Russia) and stored on the computer. A silver reference electrode was located in the chamber floor.
In present study we recorded and analyzed the changes amplitudes the both of AMPA and NMDA EPSP. Amplitudes of these FP postsynaptic components we estimated from the isoline to the peak level as shown in Figure 1A (the top row of records). The amplitudes of AMPA EPSP we assessed within an 2 msec window centered at the peak of the response. Peak NMDA EPSP was measured as the average potential observed in an 8 msec window [15].
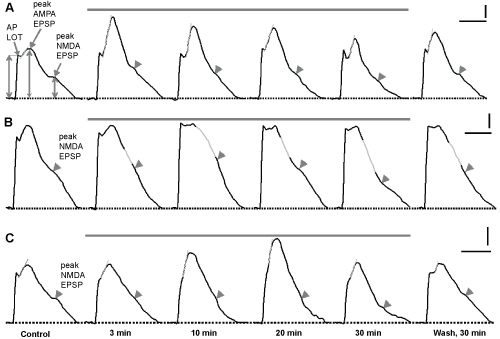 Figure 1: Representative traces of synaptic responses recorded in piriform cortex of olfactory slices at perfusion CPZ with different concentrations. FPs at perfusion with CPZ in concentrations 10-6 M (A), 10-5 M (B) and 10-4 M (C). The gray solid line in A, B and C - the duration of application CPZ. Horizontal dotted line - isoline. At the control FPs in A arrows indicate the separate components of FPs. Gray vertical arrows from the isoline to AMPA and NMDA of peaks of EPSP indicate methods of measurement the amplitudes of these components FPs. In A and C, the tangent gray lines represent the lines slope of the rising phase of AMPA EPSP. In B, a gray inclined lines indicates the slope of the descending phase AMPA EPSP. These labels are presented for estimation the degree of activation of sodium (A,C) and potassium channels (B). Numerals on the bottom of registration traces in C - time points of FPs registration. Thick gray arrows on the descending phase of the FPs indicate the peaks of NMDA EPSP. Calibration in A, B and C: 0.1 mV, 5 msec. View Figure 1
Figure 1: Representative traces of synaptic responses recorded in piriform cortex of olfactory slices at perfusion CPZ with different concentrations. FPs at perfusion with CPZ in concentrations 10-6 M (A), 10-5 M (B) and 10-4 M (C). The gray solid line in A, B and C - the duration of application CPZ. Horizontal dotted line - isoline. At the control FPs in A arrows indicate the separate components of FPs. Gray vertical arrows from the isoline to AMPA and NMDA of peaks of EPSP indicate methods of measurement the amplitudes of these components FPs. In A and C, the tangent gray lines represent the lines slope of the rising phase of AMPA EPSP. In B, a gray inclined lines indicates the slope of the descending phase AMPA EPSP. These labels are presented for estimation the degree of activation of sodium (A,C) and potassium channels (B). Numerals on the bottom of registration traces in C - time points of FPs registration. Thick gray arrows on the descending phase of the FPs indicate the peaks of NMDA EPSP. Calibration in A, B and C: 0.1 mV, 5 msec. View Figure 1
Chemical compounds for the preparation of aCSF and NH4Cl were supplied by Chimreactiv Company (Russia), Chlorpromazine hydrochloride was received from Moscow endocrine factory (Russia). CPZ was dissolved in modified aCSF, in which NaHCO3 was eliminated because it forms insoluble precipitates upon contact with CPZ was substituted for an equal amount of NaCl.
We studied the effects of the CPZ at concentrations 10-6, 10-5 and 10-4 M. These concentrations of CPZ were selected based at concentrations similar to those attained in the brain of psychotic patients [16]. At first, the slices perfused by control aCSF and FPs recorded during 15 min. Among the components of the FPs were analyzed the amplitudes of postsynaptic components, AMPA and NMDA EPSP which reflect the activities of AMPAR and NMDAR, respectively. These values considered as control for subsequent actions of CPZ. Then slices perfused CPZ in one of the above concentrations for 30 min. During this time, the both AMPA and NMDA EPSPs were recorded and analyzed. After the end of the perfusion, slices were washed by control aCSF during 30 min, and determined the activity of these receptors. FPs recorded under control conditions and in the presence of CPZ (30 min). The LOT was stimulated under control conditions and during treatment with the test substances with frequency 0.0033 Hz.
The statistical analyses of the changes in amplitudes of separate FP components were performed using the nonparametrical U test, Wilcoxon-Mann-Witney matched pairs signed-rank test (P ≤ 0.01). The data are presented as mean ± S.E.
In order to understand how in clinical conditions the CPZ modifies the activity of ionotropic glutamate receptors, it has been applied on slices in different doses. The activity of these receptors in the slice recorded following addition of CPZ at different doses in the bathing medium. The FPs registered during 30 min of CPZ action.
As shown in Figure 1A CPZ in concentrations of 10-6 M induced initial increase followed by a decrease of the amplitudes of the both AMPA and NMDA EPSP. In order to assess the degree of interaction CPZ with AMPAR and NMDAR and the duration its effects on these receptor mechanisms we has been used the test on reversibility of CPZ action during washing. This assay for reversible CPZ action at 10-6 M concentrations showed that the effects neuroleptic were resistant. At the end of the washing the both AMPA and NMDA EPSP amplitudes exceeded the control values (Figure 1A, Wash, 30 min).
Increase concentration CPZ in the perfusion medium to 10-5 M did not caused activation of the AMPARs during washing (Figure 1B). In contrast, activation NMDARs increased (Figure 1B, 10 min) and decreased by the end of action CPZ (Figure 1B, 30 min). In the test for the reversibility of activities of the both AMPARs and NMDARs were equal to control values (Figure 1B, Wash, 30 min).
CPZ in concentration 10-4 M induced the different responses of the AMPARs and NMDARs. So, amplitude AMPA EPSP increased in the initial period action CPZ (Figure 1C, 7-20 min), and at 30 min was equal to the control value (Figure 1С, 30 min). Surprisingly, activation NMDAR did not change during 20 min but at 30 min decreased below control values. In the test for the reversibility of activation of the both AMPARs and NMDARs were higher than control values (Figure 1С, Wash, 30 min).
The findings revealed "nonlinear" effects of CPZ on activity the both AMPAR and NMDAR. This indicates multiple effects of CPZ on functioning of the both AMPA and NMDA receptors.
In order to understand the effects CPZ in different concentrations on the activity of the both AMPAR and NMDAR, we studied the dose-dependent pattern. AMPA and NMDA responses were studied depending on the time of action CPZ in different concentrations as well as in a test for reversibility at washing. It has been found that when the lowest concentration CPZ (10-6 M) activation AMPAR increased. Moreover, these changes were phase: The initial transient increase in amplitude AMPA EPSP (1-7 min). At the end of the action CPZ, activation AMPAR reduced and was not statistically different from control values. In the test of reversibility at washing was discovered that activation AMPAR remained elevated and was significantly different from control values (Figure 2A). The concentration of CPZ (10-5 M) did not lead to significant changes in AMPAR both in action and in washing (Figure 2A).
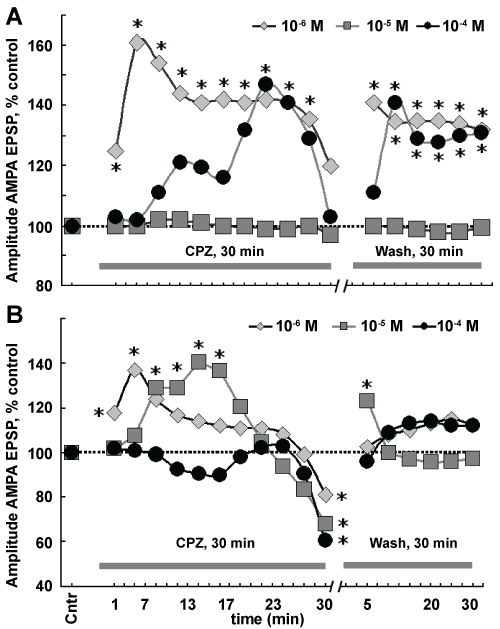 Figure 2: Effects treatment of slices with different concentrations of CPZ on the amplitudes of AMPA (A) and NMDA (B) EPSP as an index of activities AMPAR and NMDAR. Horizontal dotted line - control level for AMPA (A) and NMDA EPSP (B). The gray solid line in A and B marked "CPZ, 30 min" indicate time applications on slices in different concentrations presented in the top of the charts. The gray solid line in A and B marked "Wash, 30 min", time washout of slices by control aCSF. Abscissa - Cntr, control, the control value of AMPA (A) and NMDA EPSP (B) amplitudes referred to as control values. Numbers indicate time (min) registration during action CPZ. Wash, 30 min bathing by control aCSF. Scale time is irregular. Ordinate - designation is indicated in Figure. *P ≤ 0.05 compared with control values, nonparametric Wilcoxon-Mann-Whitney U test. n = 9 for each timepoint. SD is ommited. Detailed explanations in the text. View Figure 2
Figure 2: Effects treatment of slices with different concentrations of CPZ on the amplitudes of AMPA (A) and NMDA (B) EPSP as an index of activities AMPAR and NMDAR. Horizontal dotted line - control level for AMPA (A) and NMDA EPSP (B). The gray solid line in A and B marked "CPZ, 30 min" indicate time applications on slices in different concentrations presented in the top of the charts. The gray solid line in A and B marked "Wash, 30 min", time washout of slices by control aCSF. Abscissa - Cntr, control, the control value of AMPA (A) and NMDA EPSP (B) amplitudes referred to as control values. Numbers indicate time (min) registration during action CPZ. Wash, 30 min bathing by control aCSF. Scale time is irregular. Ordinate - designation is indicated in Figure. *P ≤ 0.05 compared with control values, nonparametric Wilcoxon-Mann-Whitney U test. n = 9 for each timepoint. SD is ommited. Detailed explanations in the text. View Figure 2
Application on slices CPZ in higher concentration (10-4 M) caused stepwise increase activation AMPAR. Small increase in amplitude AMPA EPSP was in the time range 7-15 min. Increase in activation AMPAR was significant from 16-20 min. However, further action CPZ in this concentration level of activation AMPAR declined up to control values at the end of his action. In the test for reversibility of the activation of AMPAR increased and persisted for washing (Figure 2A).
The pattern of changes of NMDA responses by the action CPZ at different concentrations differed from AMPA responses. When using the low concentrations CPZ (10-6 M and 10-5 M) was increase of activity NMDAR. Moreover, at a lower concentration CPZ (10-6 M), this was short-term excitation at 3-7 min, and then the curve was parallel to the abscissa. At both concentrations CPZ (10-6 M, 10-5 M) activation NMDAR at the end of his actions dropped significantly compared to the control. In washing NMDAR, activation was increased, but the short-term at the 5th min. While continuing to washing AMPAR activity did not differ from control values (Figure 2B).
The highest concentration CPZ (10-4 M) caused a slight inhibition of activation NMDAR with 7-15 min and maximal inhibition was at the end of action neuroleptic drug. In the test of reversibility at washing activation NMDAR increased, but these values were not statistically significant (Figure 2 B).
To further, clarify the dose-dependent characteristics CPZ action we determined these summary characteristics for fixed time point, 15 min action CPZ. This was made in order to isolate the direct effect of CPZ on AMPA and NMDA dependent mechanisms. Proportional dose effects were observed only for NMDAR, while for AMPAR such regularity was not detected (Figure 3A and Figure 3B). Probably, CPZ modifies activities separate subunits of the AMPA and NMDA receptor complexes.
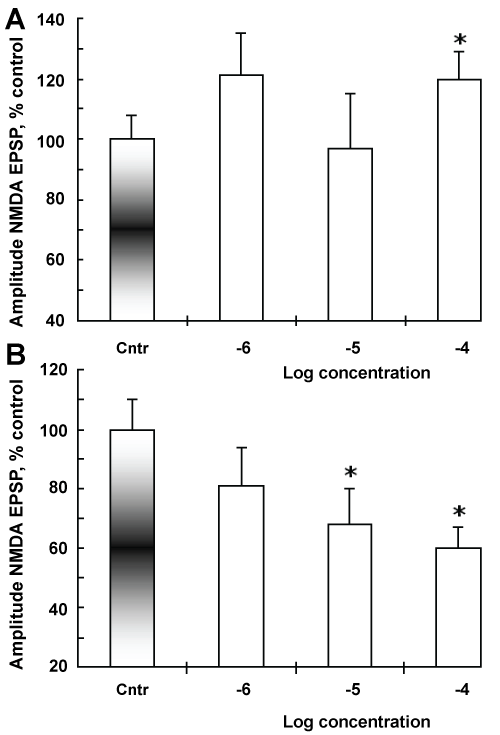 Figure 3: Dose-response relationship for changes in the amplitudes of AMPA (A) and NMDA EPSP (B) under the action of different concentrations of CPZ. Abscissa - Cntr, control values amplitude of AMPA (A) and NMDA EPSP (B); average values obtained within 15 min of perfusion slices control aCSF. The numbers, log concentrations of CPZ. Ordinate - designation is indicated in Figure. *P ≤ 0.05 compared with control values, nonparametric Wilcoxon-Mann-Whitney U test. n = 6 for each timepoint. Note, that proportional the dose-dependence was obtained only for the activity of NMDAR, whereas the rectilinear dose-dependence for AMPAR activity was not detected. View Figure 3
Figure 3: Dose-response relationship for changes in the amplitudes of AMPA (A) and NMDA EPSP (B) under the action of different concentrations of CPZ. Abscissa - Cntr, control values amplitude of AMPA (A) and NMDA EPSP (B); average values obtained within 15 min of perfusion slices control aCSF. The numbers, log concentrations of CPZ. Ordinate - designation is indicated in Figure. *P ≤ 0.05 compared with control values, nonparametric Wilcoxon-Mann-Whitney U test. n = 6 for each timepoint. Note, that proportional the dose-dependence was obtained only for the activity of NMDAR, whereas the rectilinear dose-dependence for AMPAR activity was not detected. View Figure 3
Whether it is possible, to eliminate the negative effects of CPZ on ionotropic glutamatergic receptors that have been revealed and described above? To test this hypothesis, we used pretreatment of slices using ammonium chloride (NH4Cl). NH4Cl is a very effective inhibitor of lysosomal function and oppresses the function of lysosomal proteases. This compound (Na+/H+-ionophore) is commonly recognized as fast and efficient lysosomal inhibitor [17,18]. We have suggested that ammonium chloride reduces the high lipophilicity of CPZ and prevents deterioration NMDAR functions. For the preparation of aCSF, containing ammonium chloride, 20 mM sodium chloride was removed from the incubation solution, respectively. It has been found that NH4Cl, in range 10-30 mM did not alter the activity of NMDAR (control, 73.1 ± 3.6% vice versa, NH4Cl, 70.9 ± 3.7%, nonparametric Wilcoxon-Mann-Whitney U = 23, n = 7, P > 0.05).
Incubation slices with NH4Cl and subsequent action of CPZ (10-5 M) shifted the pattern of NMDAR responses (Figure 4). The initial activation of these receptors has been short-term (7.1 ± 2.3 min). Peak amplitudes changes of NMDA EPSP did not differ from control values (NH4Cl, 20 mM + CPZ, 10-5 M: U = 25; n = 7; P ≥ 0.05 vice versa CPZ, 10-5 M: U = 8; n = 7; P ≤ 0.05). Period reducing activity of NMDAR was gradual. At washing activity of these receptor mechanisms remained unchanged (Figure 4). These findings indicate that CPZ alters phasic manner the functioning of NMDAR. Importantly, lysosomal inhibitor NH4Cl eliminates the negative effects CPZ on the activity of NMDAR. We believe that the recovery of NMDAR activity was because the NH4Cl blocked the high lipophilicity CPZ and eliminated its negative effects on these receptors.
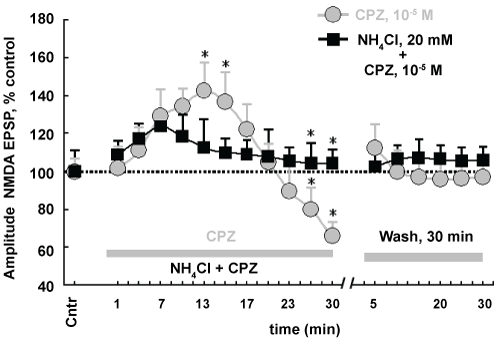 Figure 4: Effects of lipophilicity CPZ on NMDAR activity. Horizontal dotted line - control level of NMDA EPSP amplitude. On top gray solid line marked "CPZ" is shown time of applications CPZ on slices in concentration 10-5 M during 30 min. Lower gray solid line marked "NH4Cl + CPZ" indicates pretreatment of slices by "lysosomal inhibitor" NH4Cl (20 mM) during 15 min and subsequent action of CPZ (10-5 M) during 30 min. The gray solid line marked "Wash, 30 min", time washing of slices by control aCSF. Abscissa - Cntr, control, the control value of NMDA EPSP amplitudes. Numbers indicate time (min) registration of NMDA EPSP during action NH4Cl, CPZ. Wash, 30 min bathing slices by control aCSF. Scale time is irregular. Ordinate - designation is indicated in Figure. *P ≤ 0.05 when comparing a data on curve "CPZ, 10-5 M" with the data on the curve "NH4Cl, 20 mM + CPZ, 10-5 M", nonparametric Wilcoxon-Mann-Whitney U test. n = 7 for each time point. View Figure 4
Figure 4: Effects of lipophilicity CPZ on NMDAR activity. Horizontal dotted line - control level of NMDA EPSP amplitude. On top gray solid line marked "CPZ" is shown time of applications CPZ on slices in concentration 10-5 M during 30 min. Lower gray solid line marked "NH4Cl + CPZ" indicates pretreatment of slices by "lysosomal inhibitor" NH4Cl (20 mM) during 15 min and subsequent action of CPZ (10-5 M) during 30 min. The gray solid line marked "Wash, 30 min", time washing of slices by control aCSF. Abscissa - Cntr, control, the control value of NMDA EPSP amplitudes. Numbers indicate time (min) registration of NMDA EPSP during action NH4Cl, CPZ. Wash, 30 min bathing slices by control aCSF. Scale time is irregular. Ordinate - designation is indicated in Figure. *P ≤ 0.05 when comparing a data on curve "CPZ, 10-5 M" with the data on the curve "NH4Cl, 20 mM + CPZ, 10-5 M", nonparametric Wilcoxon-Mann-Whitney U test. n = 7 for each time point. View Figure 4
In the present study we investigated the effects of CPZ, a widely used antipsychotic, on the activity of ionotropic glutamatergic receptors, AMPAR and NMDAR. We found that CPZ interacts with both ionotropic glutamate receptors AMPAR and NMDAR. CPZ demonstrated expressed neurotrophic effects on these receptors mechanisms: It modified the amplitude/temporal characteristics of the activity of these receptors and altered the kinetics of the complicated interactions with these receptor complexes.
The effects of CPZ on activity the both AMPAR and NMDAR were studied in the in brain slices technique on rat olfactory neurons, recording extracellular field potentials in the piriform cortex of the olfactory brain slices.
The phenothiazine and its derivatives are characterized by low ionization potentials and the active radicals are formed, it react with many substances such as proteins, alcohol, carbonates, and many drugs [4]. These data were taken into account in our research when studying the effects of CPZ on the both AMPA and NMDA receptors in brain slices. Thus, carbonate salts, proteins and peptide neuroprotectors were excluded from the incubation solution.
Moreover, electrophysiological analysis indicated that, at concentrations similar to those found in the cerebrospinal fluid of schizophrenics, when assessing dose-relationships, CPZ nonlinear way modified AMPAR activity with increasing concentrations. Opposite NMDAR activity decreased in a linear manner with increasing concentrations CPZ in incubation medium.
The nonlinearity of CPZ effects can be explained by the formation of active radicals. Reactions of these substances with neurons and their membranes have not yet been established. Perhaps their formation contributes to decrease of lipophilicity, and this process was recorded by us in the reduction of the amplitude of NMDA EPSP after 20 min of CPZ action (Figure 2).
In the explanation of these findings should be based on facts that CPZ modified the physicochemical properties of the membranes of neurons. In our opinion, it is due to high lipophilicity CPZ. So, CPZ increased membrane fluidity and permeability of the neuroleptic into neurons. It is result in overall destabilization of cell membranes. It was revealed that the distribution of aliphatic-type phenothiazine-derivate neuroleptic drugs depends largely on their interaction with phospholipid membranes [19]. Important, that CPZ has a positive charge, making it actively interacting with negatively charged cell membrane lipids [20]. It is established also that CPZ caused the formation of 14 Å diameter holes in the membranes of red cells. These data cogently demonstrate CPZ can modify membrane of neurons and accordingly change sodium and potassium currents involved in the generation of AMPA EPSP.
With prolonged action CPZ modification activities AMPAR and NMDAR at different concentrations was phase: Initial activation and subsequent depression. However, it has been found in our experiments for AMPAR at concentrations of CPZ 10-6 M and 10-4 M. As for the NMDAR at low concentrations, CPZ (10-6 M and 10-5 M) was the initial activation, and at the end of exposure, these receptors were depressed. In the test for the reversibility of effects, CPZ at washing initial activity of both AMPAR and NMDAR restored.
We suggest that pattern of activation of these receptors is associated with the effect of CPZ on different sites of AMPAR and NMDAR. We believe that inhibitory effect of CPZ can be explained the protective effects this antipsychotic, which detected in the different models. The protective effects of CPZ on the nervous system are supported the both in vitro and animal studies including hypoxia [21], ischemia [22] and glutamate-induced neurotoxicity [23].
Presented above data indicate that CPZ at long action negatively affects on ionotropic glutamatergic receptor mechanisms. To eliminate these adverse effects CPZ we took into consideration, that CPZ has a high lipophilicity. This antipsychotic increased membrane fluidity and permeability neurons membranes. It was revealed that the distribution of aliphatic-type phenothiazine-derivate neuroleptic drugs, including CPZ, depends largely on their interaction with phospholipid membranes [19]. This leads to changes in membrane structure and its functions.
It is revealed, CPZ penetrates into cells and promotes penetration of some substances: ATP, ADP, glucose 6-phosphate, and fructose 1,6-bisphosphate [24,25]. Perhaps, CPZ penetration into neurons reduces NMDA EPSP amplitude after 20 min of neuroleptic action (Figure 2A). We suppose that NMDA channels may be port for penetration CPZ molecules into cells. At the same time, this complex issue requires further research.
To solve the issue negative effects of CPZ on the both AMPA and NMDA-dependent mechanisms the experiment was carried out in the presence of "lysosomal inhibitor", ammonium chloride. This compound (Na+/H+-ionophore) is commonly recognized as fast and efficient lysosomal inhibitor [18]. Lysosomal trapping is an important mechanism of cellular and body distribution of basic psychotropic drugs, such as phenothiazine neuroleptics [19]. We hypothesised that ammonium chloride reduces the high lipophilicity of CPZ and protects deterioration NMDAR functions.
Our experimental findings indicate that NH4Cl eliminated depressive effects CPZ on activity NMDARs. It should be noted pretreatment of slices by NH4Cl did not affect the interaction of CPZ with glutamate site NMDAR, since the initial NMDAR activation was reduced but short-term. However, NH4Cl inhibited the late activation NMDARs induced by CPZ. In the test of reversibility (in the clinical sense, this means the termination actions of CPZ on neurons). It was found that pretreatment of slices by NH4Cl led to complete recovery of NMDAR activity. These findings indicate that inhibitor lipophilicity NH4Cl eliminate negative CPZ effects on NMDAR and optimize their functioning.
Presented data extend the understanding a range of the CPZ effects in the CNS. Currently, the dominant idea is that the effects of CPZ are mainly manifested through dopamine and serotoninergic receptors. However, our data indicate that CPZ actively interacts with ionotropic glutamatergic mechanisms. Note that the glutamatergic system is the main excitatory mediator system in the CNS mammals, including human. These data prove that the glutamatergic system is actively involved in schizophrenia. We believe data presented in this study on the influence of CPZ on the ionotropic glutamatergic mechanisms allow understanding the multifaceted mechanisms action of neuroleptics in preventing the deterioration of mental disorders such as schizophrenia.
None declared.
The author thanks Mrs. Galina Smirnova for her excellent technical assistance.