Skin cancer is the most common form of cancer in the United States of America and in Australia. Basal cell carcinoma (BCC), the most prominent skin cancer, is curable, but can be disfiguring and costly to treat.
Treatment of BCC is important, because untreated tumours can grow incessantly and invade underlying tissue. Surgery has the highest cure rate for BCC. Other effective treatments for BCC include radiation therapy, laser therapy, electrosurgery, cryotherapy, fluorouracil and imiquimod therapies. All of these skin cancer therapies are indiscriminate with necrotic, unregulated, chaotic cell death sequelae.
Lack of selectivity and specificity with resulting necrosis restrict these therapies, as evidenced by scarring, recurrence of the treated lesion and limited cosmetic outcomes. Curaderm is a selective drug that has the ability to discriminate between skin cancer cells and normal skin cells. Furthermore, Curaderm is highly specific and is capable of causing cancer cell death by the orderly process of apoptosis.
The clinical aspects of Curaderm pharmacotherapy have amply been described in the literature. Recently, a thought-provoking article "Curaderm, the long-awaited breakthrough for basal cell carcinoma" was published. Although the article is awe- inspiring, it does not address the mechanisms and mode of actions of Curaderm that achieve such remarkable clinical outcomes.
This communication centres on the pharmacological mechanisms, which establish that Curaderm is indeed both selective and specific in eliminating basal cell carcinoma from the skin.
Cancer, Curaderm, BEC, Solamargine, Apoptosis, BCC, Basal cell carcinoma, Skin cancer
BCC is the most common form of skin cancer and the most frequently occurring form of all cancers in humans. An estimated 3.6 million cases are diagnosed each year in the U.S. and this number is steadily increasing.
BCC most often occurs when DNA damage from over-exposure to ultraviolet (UV) radiation from the sun triggers undesirable changes in the basal cells in the outermost layer of the skin. This can result in uncontrolled growth of the affected basal cells leading to BCC.
Treatments of BCC may involve:
• Surgical removal consisting of excision, Mohs surgery or curettage and electrodesiccation
• Cryotherapy
• Radiation
• Photodynamic therapy
• Imiquimod
• 5-Fluorouracil
When found early and treated by the above procedures, the BCC can often be removed. However, depending on the procedure used, because of the non-selectivity and non-specificity of the treatment procedure, the recurrence rate of the BCC can be high and the cosmetic outcome suboptimal.
Because of lack of selectivity and specificity, virtually all of the above treatments are indiscriminate in their actions and result in necrosis of cancer cells, including normal cells.
Curaderm is a topical treatment for BCC that contains natural allelochemical secondary metabolic glycoalkaloids with robust anticancer properties [1,2].
Curaderm contains BEC, which is a mixture of tri-, di-, and mono-glycosides of solasodine. The triglycoside solamargine is the most effective anticancer component and represents 86% of total anticancer activity [1,3,4]. Solamargine, a small-molecule drug, is a triglycoside of which the alkaloid solasodine is conjugated with two molecules of the plant sugar rhamnose and one molecule glucose.
Cell culture, animal and human studies have determined the efficacy of BEC and its individual components with high therapeutic indices for a wide variety of cancers [1-27].
Recently, a thought-provoking review article described the clinical aspects of Curaderm, a solamargine containing topical cream treatment for BCC [1].
However, the pharmacology of Curaderm therapy was not addressed in the review article. To fully appreciate the safety, high efficacy and remarkable cosmetic results obtained with Curaderm therapy, it is important to understand the pharmacological aspects of selectivity, specificity and mode of cancer cell death.
Solamargine in Curaderm, through its rhamnose content (Figure 1 highlighted area) selectively identifies and binds to cancer cells.
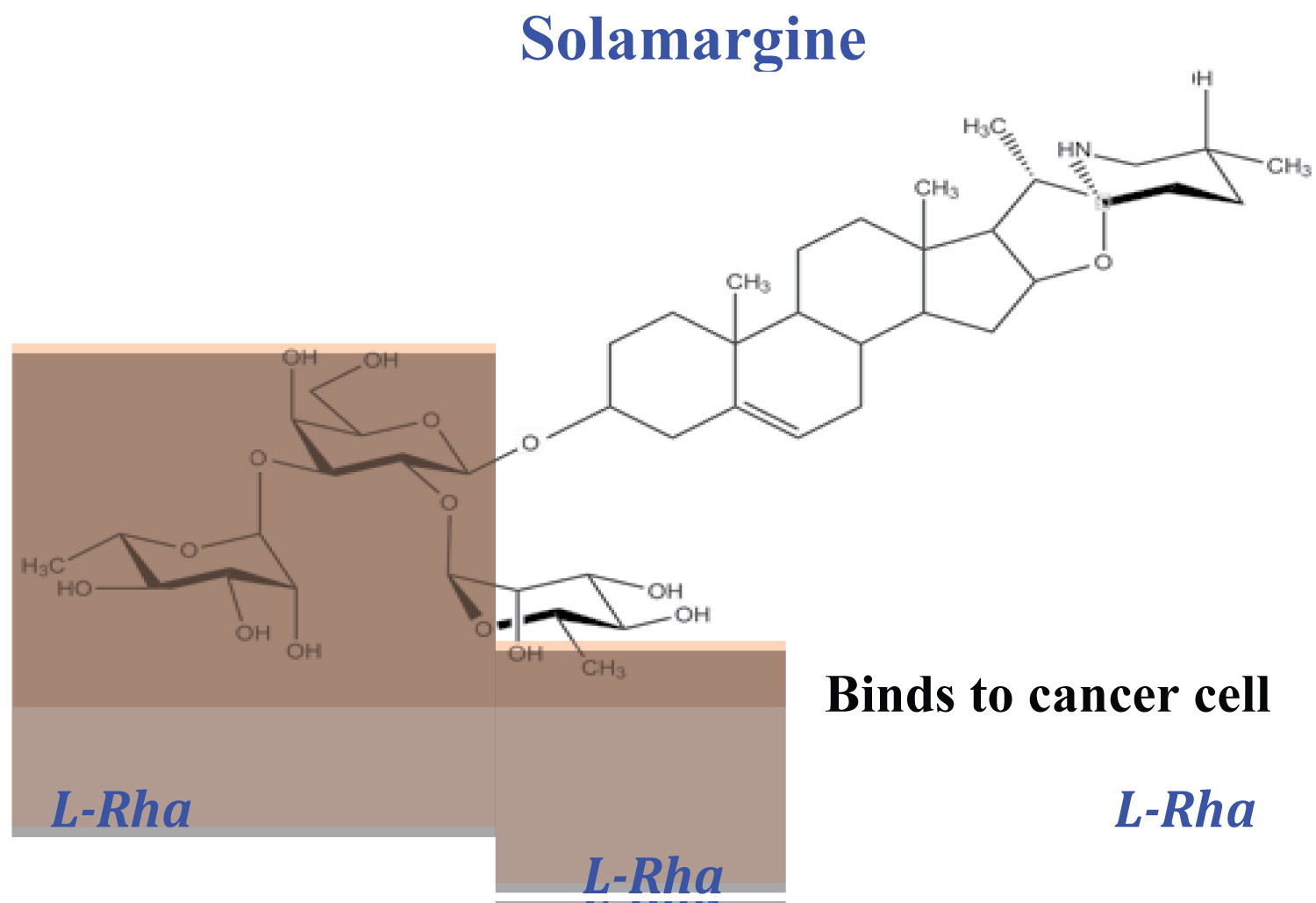 Figure 1: Chemical structure of solamargine, the main anticancer ingredient in Curaderm. The rhamnose sugar moiety (highlighted area) selectively identifies and interacts with specific rhamnose binding proteins (RBP) that are located on the surface of cancer cells.
View Figure 1
Figure 1: Chemical structure of solamargine, the main anticancer ingredient in Curaderm. The rhamnose sugar moiety (highlighted area) selectively identifies and interacts with specific rhamnose binding proteins (RBP) that are located on the surface of cancer cells.
View Figure 1
Cell culture and whole animal studies show conclusively that cancer cells contain specific endogenous endocytic lectins (EELs) [1-5]. These lectins have been identified and characterised as rhamnose binding proteins (RBP) and are located on the surface of cancer cells, whereas normal non-cancer cells lack these receptors. Rhamnose is a plant sugar and is normally not present in the human body.
RBP on cancer cells are mutant proteins that possess cancer cell-specific binding affinities with rhamnose and its conjugates. The number of RBP receptors on the various cancer cell types range from 500-10,000 per cell with an affinity constant of 1.5 x 10-5 M and a molecular weight of 67 kDa [4,6]. Different cell populations of specific cancers establish that a single type of RBP receptor is involved in virtually all cancer cell lines. There are two distinct binding categories of solamargine to cancer cells, which are related to receptor affinity and numbers of receptors per cell [4,6,28,29]. Cutaneous BCC, squamous cell carcinoma and malignant melanoma possess receptors with high affinity for solamargine.
Cellular, animal- and human cancer cell competitive inhibition studies of BEC with free, unbound rhamnose have established that free, unbound rhamnose interferes with the efficacy of solamargine. It is therefore essential to ensure the absence of free, unbound rhamnose in the BEC-containing formulations [1-5].
Fluorescence imaging analysis of a synthesized biotinylated-rhamnose probe has visualized the expression of RBP receptors on cancer cells but not normal cells (Figure 2) [29].
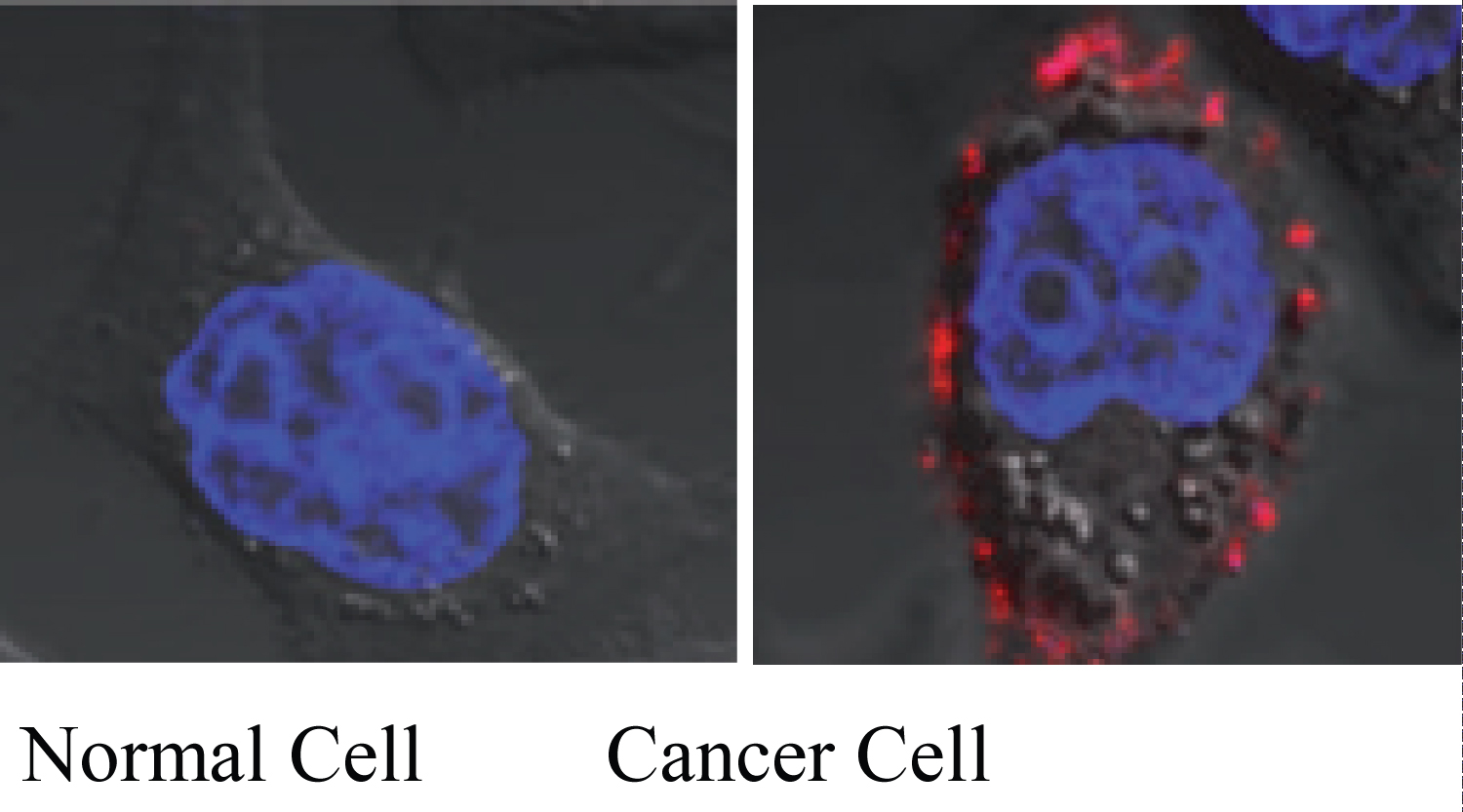 Figure 2: Identification of specific rhamnose binding protein (RBP) receptors on cancer cells by a fluorescence rhamnose probe. Normal non-cancer cells do not possess the RBP receptors. The red images indicate the binding of rhamnose conjugates (solamargine) to RBP receptors.
View Figure 2
Figure 2: Identification of specific rhamnose binding protein (RBP) receptors on cancer cells by a fluorescence rhamnose probe. Normal non-cancer cells do not possess the RBP receptors. The red images indicate the binding of rhamnose conjugates (solamargine) to RBP receptors.
View Figure 2
Using this technique, it was shown that very low concentrations of free rhamnose completely blocked the binding of the probes to cancer cells (Figure 3) [2-6] [29,30].
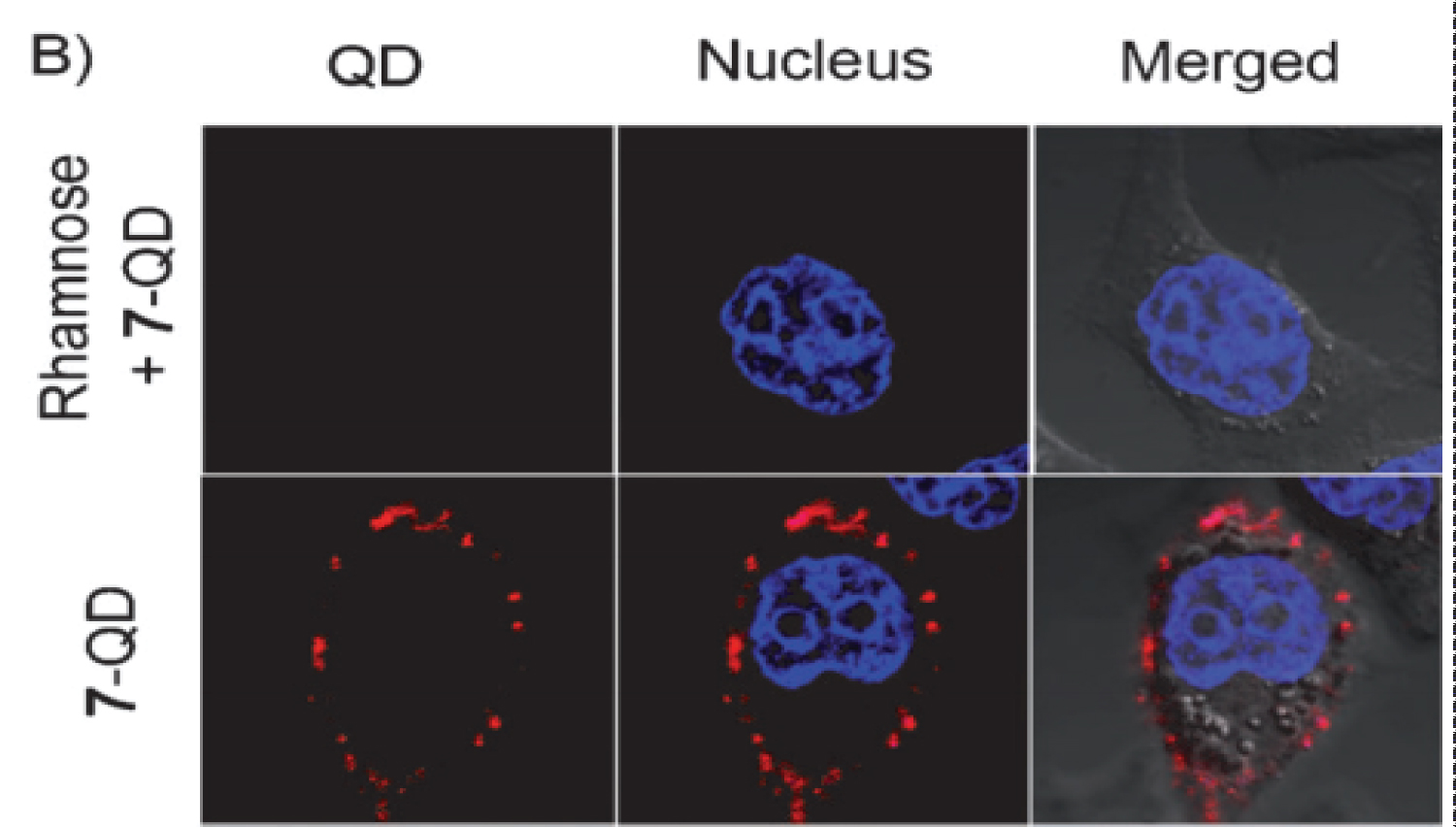 Figure 3: Confocal microscopic view of rhamnose-QD probe (representing solamargine) localized on the cell surface of human squamous cell carcinoma (KB) cells. The binding of the probe was blocked by rhamnose. Confocal microscope image of KB cells labelled by probe (red) and Hoechst stain (blue).
View Figure 3
Figure 3: Confocal microscopic view of rhamnose-QD probe (representing solamargine) localized on the cell surface of human squamous cell carcinoma (KB) cells. The binding of the probe was blocked by rhamnose. Confocal microscope image of KB cells labelled by probe (red) and Hoechst stain (blue).
View Figure 3
Indeed, at a clinical level the presence of free rhamnose imparts inhibitory properties to the efficacy of BEC or solamargine formulations when treating terminal tumours and skin cancers [4].
After identification and binding of solamargine to its specific RBP receptor, solamargine is internalized into the cancer cell by receptor-mediated endocytosis through endosomes and ultimately in the lysosomes and mitochondria. Consequently, the moiety of the alkaloid solasodine (Figure 4, highlighted area) exerts apoptosis by increasing pro-apoptotic Bax and Bak proteins and decreasing the anti-apoptotic Bcl-2 and Bcl-x proteins at the transcriptional and translational phases. Caspase-9 and Caspase-3 are activated during these processes, resulting in apoptosis [2-4] [31-37].
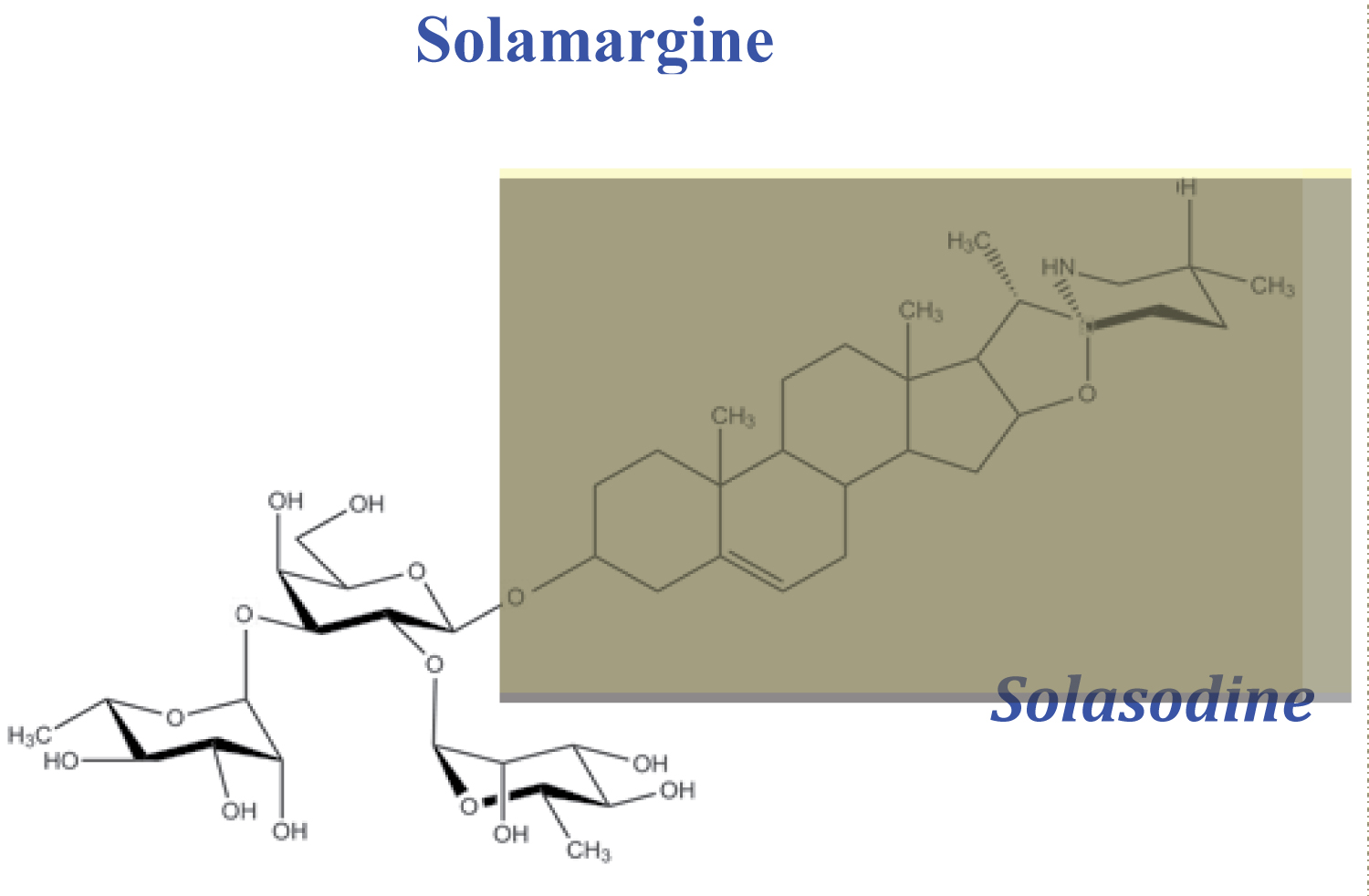 Figure 4: After specific internalisation of solamargine (SM) by receptor-mediated endocytosis into the cancer cell, the alkaloid moiety solasodine (highlighted area) exerts apoptosis in the cancer cell.
View Figure 4
Figure 4: After specific internalisation of solamargine (SM) by receptor-mediated endocytosis into the cancer cell, the alkaloid moiety solasodine (highlighted area) exerts apoptosis in the cancer cell.
View Figure 4
Solamargine and BEC also induce the extrinsic apoptotic death pathway in cancer cells by up-regulating the expressions of external death receptors such as tumour necrosis factor receptor 1 (TNFR-1) and 6 (TNFRSF6 or Fas receptor), TNFR-1-associated death domain (TRADD) and Fas-associated death domain (FADD) [2-4] [31-37]. Figure 5 illustrates schematically the process of apoptosis.
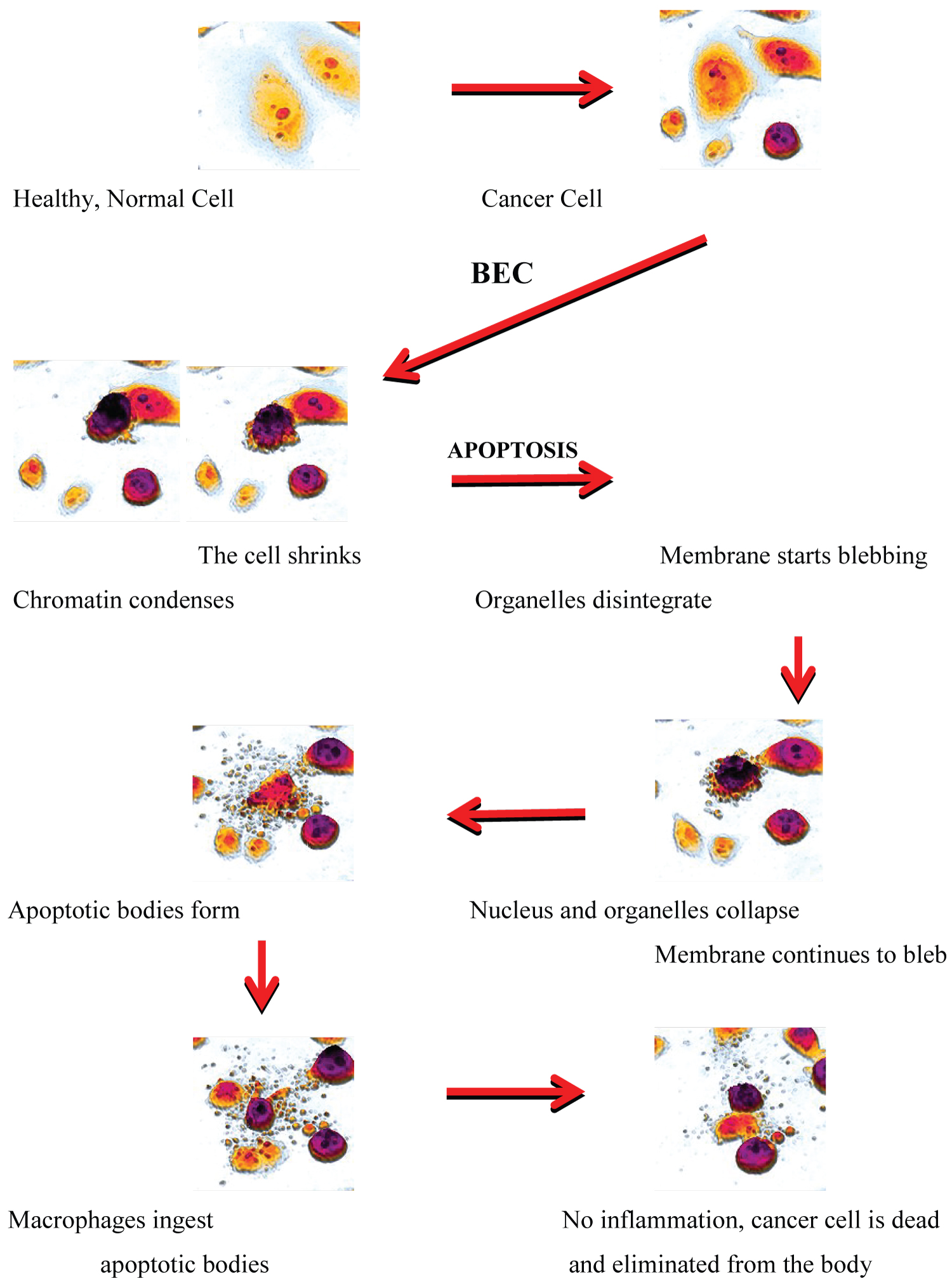 Figure 5: Schematic representation of solamargine (SM)-induced apoptosis by identifiable anti cancer properties through a variety of biological pathways in the cancer cell. The cancer cell dies neatly, without damaging its neighbours.
View Figure 5
Figure 5: Schematic representation of solamargine (SM)-induced apoptosis by identifiable anti cancer properties through a variety of biological pathways in the cancer cell. The cancer cell dies neatly, without damaging its neighbours.
View Figure 5
The impressive sensitivity of selectivity of Curaderm towards malignant cancer cells is shown by studies whereby solamargine is able to clearly discriminate primary and metastatic melanoma cells from benign melanoma cells or normal non cancer cells, a remarkable feature (Figure 6) [6].
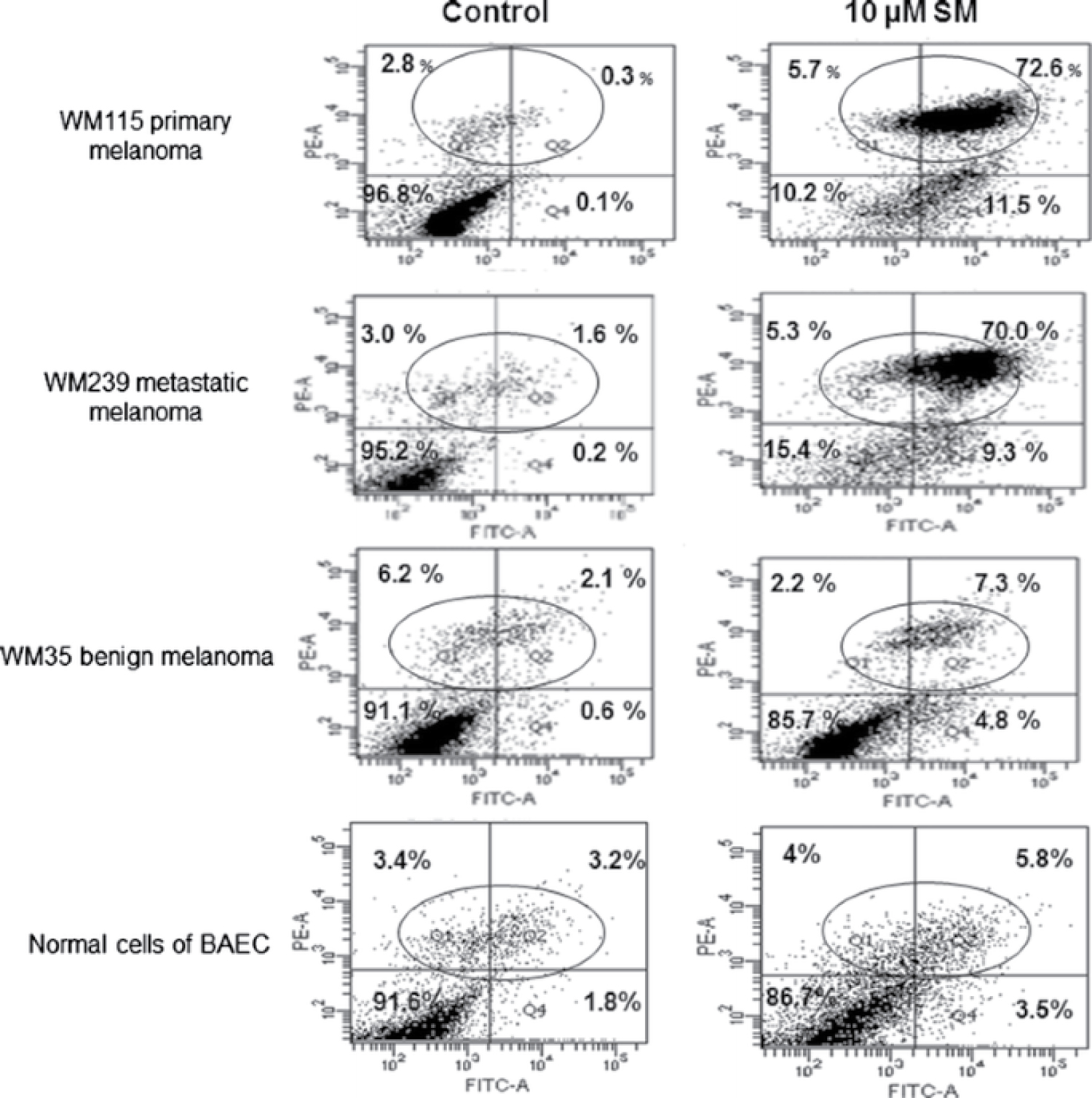 Figure 6: Flow cytometry analysis showed that solamargine (SM) increased cell death tenfold more in malignant to benign cells. 10 μM Solamargine treatment for 2 h selectively caused massive cellular death of WM115 primary and WM239 metastatic melanoma cells, which were at least tenfold, more than that detected in benign WM35 and/or in bovine aortic endothelial cells (BAEC) normal cells.
Figure 6: Flow cytometry analysis showed that solamargine (SM) increased cell death tenfold more in malignant to benign cells. 10 μM Solamargine treatment for 2 h selectively caused massive cellular death of WM115 primary and WM239 metastatic melanoma cells, which were at least tenfold, more than that detected in benign WM35 and/or in bovine aortic endothelial cells (BAEC) normal cells.
The top right quadrant of the figures indicates cell death. It is seen that solamargine (SM) kills 72.6% of primary melanoma cells and that no treatment kills only 0.3% of these cancer cells (top row). Similarly, SM kills 70% of metastatic melanoma and no treatment kills 1.6% of these cancer cells (second row). Importantly, SM does not significantly kill benign melanoma cells (untreated 2.1%, SM treated 7.3%) (Third row), nor does SM kill normal cells (untreated 3.2%, SM treated 5.8%) (Fourth row).
View Figure 6
For a drug to be able to identify an undesirable cancer cell amongst normal cells is a major feat. But, for that drug to also be able to specifically kill the undesirable cancer cell and not affect surrounding normal cells is extraordinary. This is exactly what Curaderm does when treating cutaneous BCCs!
At a clinical level compelling evidence of selectivity and specificity of BEC towards cancer cells resulting in apoptosis occurs when treating skin cancers [1-4].
The BEC-containing cream formulation Curaderm seeks and destroys BCC cells by apoptosis whilst normal skin cells replace the dead cancer cells DURING therapy. This indicates that BEC only affects cancer cells and not normal cells. Curaderm locates and eliminates cancer cells at the microscopic level and also kills cancer cells not seen with the naked eye. Visually, during Curaderm therapy, the treated skin cancer lesion appears to initially become larger until the cancer cells are eliminated.
This is followed by reduction of skin lesion until complete removal of the skin cancer cells and replacement with normal cells occurs [1-4][7]. The ratio of cancer cell death by apoptosis and new normal skin cell growth determines the size of the visual observed lesion. This phenomenon reflects self-titration, whereby Curaderm treatment only ceases when the lesion is completely healed.
It is important to appreciate that whilst skin cancer cells are undergoing apoptosis during treatment with Curaderm, regeneration of new epidermis at the application site occurs. This phenomenon is supported by the preclinical and clinical observations that Curaderm is preferential in its action towards transformed cells and explains the impressive clinical and cosmetic outcomes [1-7].
These underlying unique pharmacological properties of Curaderm culminate in apoptosis and have led to the treatment of the various types of cutaneous BCCs, including superficial, nodular and infiltrating (morpheoic) BCCs. Moreover, BCCs that are located at difficult-to-treat areas are also very amenable to Curaderm therapy [1-4][7].
The following cases (Figure 7) show that because of selectivity, specificity, efficacy, cell death by apoptosis and autophagocytosis, Curaderm therapy for BCC is as good or even better than surgical excision, the considered gold standard method, for treating BCC [38-58].
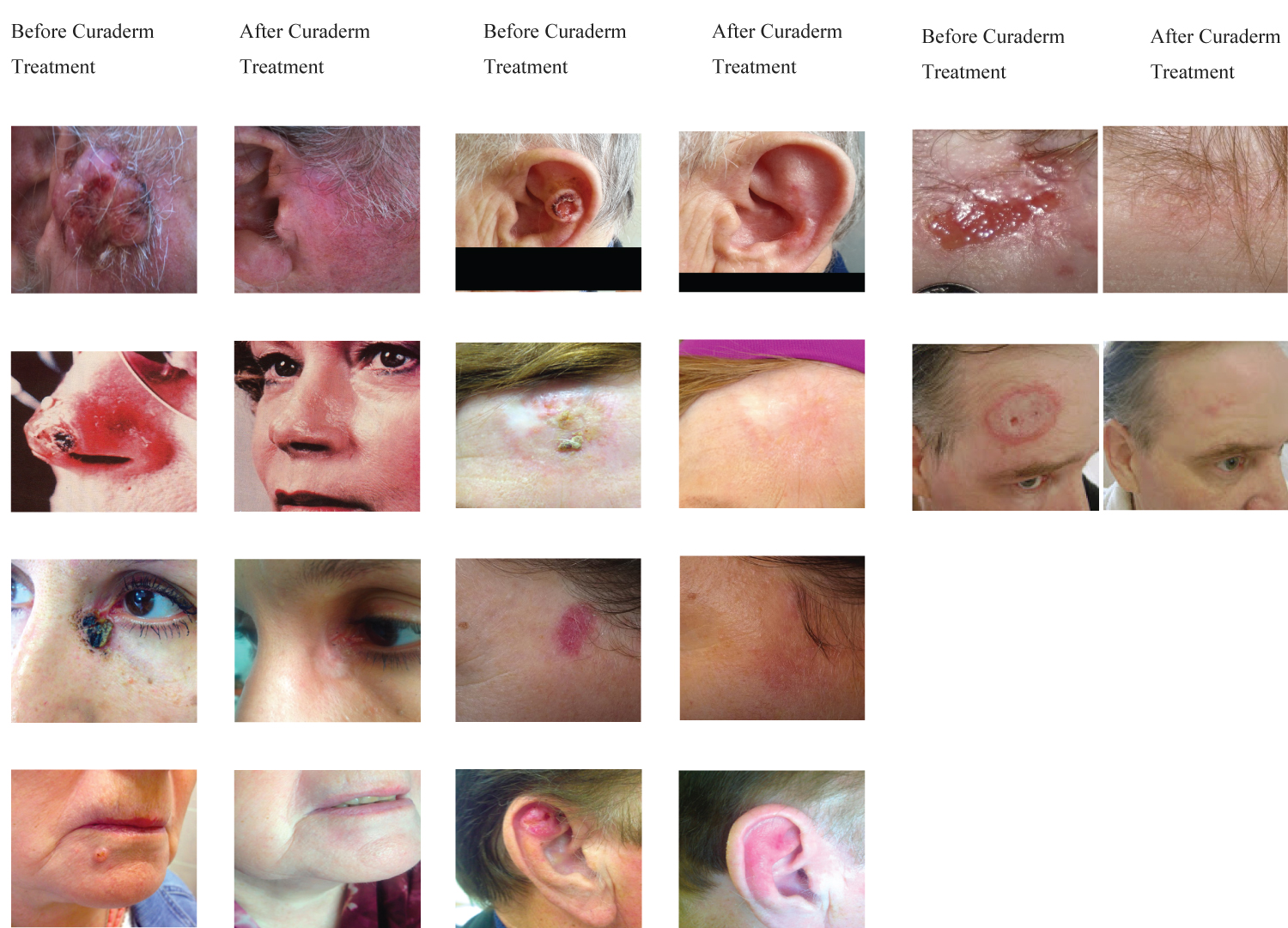 Figure 7: Examples of poor treatment prognoses of BCC lesions due to size, type, depth, previous failure by other treatment modalities and location (column, Before Curaderm Treatment) and the impressive cosmetic outcomes of Curaderm therapy by simple topical application (column, After Curaderm Treatment). Curaderm-treated lesions did not recur for at least 5 years follow-up.
View Figure 7
Figure 7: Examples of poor treatment prognoses of BCC lesions due to size, type, depth, previous failure by other treatment modalities and location (column, Before Curaderm Treatment) and the impressive cosmetic outcomes of Curaderm therapy by simple topical application (column, After Curaderm Treatment). Curaderm-treated lesions did not recur for at least 5 years follow-up.
View Figure 7
Curaderm contains 0.05 mg BEC/mL cream. Therapeutically, for large lesions less than 1 mL of Curaderm is applied to the treated lesion and two applications per day are recommended during treatment. Ripe eggplant fruits contain 20-50 mg BEC/kg fresh weight. An average eggplant fruit weighs approximately 300 g and contains approximately 15 mg of BEC.
This is more than 300 times the amount of BEC applied to the skin. In other words, the amount of BEC applied to the skin in the form of Curaderm is less than 1gram of extracted eggplant fruit!!!
Additionally, it was shown that 1 mg BEC/kg human body weight is safely administered intravenously into humans [2-8]. This translates to a person weighing 60 kg being able to tolerate 60 mg BEC in their blood.
This compares to a dose of BEC-1200 times more than is applied to the skin with Curaderm.
BEC concentrations in human plasma of 4000 ng/mL have been reported to be tolerable [8]. In Phases I-III using different concentrations of BEC in various cream formulations, no BEC could be detected in the plasma of the patients. The sensitivity of the HPLC-mass spectral analysis of BEC was 1 ng/mL plasma. This indicates that BEC in the cream formulations were safe by a factor of over 4000 [4].
All other ingredients in Curaderm, including salicylic acid and urea, have been, and are currently being used worldwide at much higher concentrations in over-the-counter (OTC) skin care products. The safety of the entire Curaderm formulation, including all ingredients, has been supported by biocompatibility studies [37].
A wide range of toxicological and pharmacological studies of BEC, including toxicokinetics, pharmacokinetics and pharmacodynamics, confirm the high efficacy and safety of Curaderm [2-5].
Curaderm therapy selectively, specifically, effectively and safely eliminates all types of BCC skin cancers by a process of apoptosis, culminating in excellent cosmetic outcomes. Curaderm is an ideal therapy for outpatient treatment.
Its limitation is the duration of therapy, which is dependent on the size, type, and location of the BCC. Transient local irritation due to the keratolytic agents in the formulation is mild to moderate, nevertheless, compliance is high and Curaderm is well tolerated. Recurrences of Curaderm-treated BCCs are very low. Curaderm has been available and is used by the public for over two decades with no reported serious adverse events. Curaderm is a thoroughly researched product and should be considered as a major additional treatment for BCC.