Despite being traditionally used herbals to treat mild anxiety and sleep disorders, Valeriana officinalis and Melissa officinalis mechanism of action is not fully understood. While the pattern of mood modulation of both herbals suggests the involvement of the Gamma-Aminobutyric Acid (GABA) ergic system, other changes in the neurotransmitters pathways can provide an explanation for the clinical effects. The aim of this study was to examine the combined mechanism of action of the two extracts in order to explain the anxiolytic and calming effects that may facilitate sleep induction and improve sleep quality in patients with mild to moderate sleep disturbances. These effects have been previously reported in the literature for a botanical product combining 120 mg Valeriana officinalis and 80 mg of Melissa officinalis standardized extracts (Songha® Night). To assess mechanism of action we employed the Unpredictable Chronic Mild Stress (UCMS) animal model, as it has strong predictive validity. Brain GABA and glutamate levels were sampled using a microdialysis technique and measured by HPLC. Additional in vitro experiments investigated the effects of the extracts on Group II Metabotropic Glutamate Receptors (mGlu2) binding and GABA Transaminase (GABA-T) activity. Results indicate that the two extracts together exert a functional synergism leading to normalization of brain levels of GABA and glutamate altered by stress. In vitro experiments reveal that the combination of both extracts could inhibit GABA-T activity more effectively than the single extracts and that Valeriana officinalis binds mGlu2 receptors. Altogether these results demonstrate a synergistic effect of Valeriana officinalis and Melissa officinalis standardized extracts on counteracting the alterations in neurotransmitter levels caused by chronic stress and support the tranquilizing and sleep inducing activity of Songha® Night observed in clinical trials.
Valeriana officinalis, Melissa officinalis, GABA, Glutamate, Sleep
Sleep loss and sleep disorders are among the most common health problems. Surveys report that approximately 30% to 40% of adults have problems initiating or maintaining sleep [1-3]. About 10% to 15% of adults report severe problems and this statistic rises up to 25% in the elderly [2]. Insomnia is found in up to 69% of patients attending primary care clinics and more common with chronic medical conditions [3,4]. Chronic sleep loss has serious consequences for health, performance, and safety [5]. Anxiety and stress disorders represent a range of conditions that include generalized anxiety, panic attacks, post-traumatic stress disorder, obsessive-compulsive syndrome, and social phobias. All of them lead to sleep disturbances and interfere with cognitive functions [6]. Unfortunately, there is only limited evidence to support the efficacy of many of the commonly used remedies for insomnia, including antihistamines, chloral hydrate, barbiturates, tryptophan and melatonin [7]. Although benzodiazepines are known to be effective for insomnia, their clinical benefit is negligible and similar to that found with exercise therapy alone [8]. Moreover, benzodiazepine receptor agonists chronic treatment is limited by concerns regarding long-term efficacy and the potential for abuse, dependence, and adverse side effects, including an increased risk of motor vehicle accidents, falls and fractures [7]. For these reasons, many people with sleep disorders resort to complementary medicines, including herbal extracts [9].
Two herbal extracts, Valeriana officinalis (VAL) and Melissa officinalis (MEL) have been commonly used to treat mild anxiety and sleep disorders [10,11]. The extract of the root of VAL, a flowering plant, has been widely used to treat sleep disorders in Europe for decades [10] and is becoming increasingly popular in the United States as a self-prescribed treatment for insomnia. In a national survey, approximately two million adults reported using valerian in the past week [11].
The perennial lemon-scented herb MEL (synonym lemon balm) has been in use as a pan-cultural medicinal treatment for more than two millennia. Its traditional indications have included general beneficial effects on the brain as a treatment for memory disorders [12] and for “all complaints supposed to proceed from a disordered state of the nervous system” [13]. Contemporary reports emphasize the sedative, spasmolytic, and antibacterial effects of MEL, with indications encompassing nervous, gastrointestinal, and sleep disorders [14-16].
The specific mechanisms of action responsible for the pharmacological effects of VAL and MEL have not been fully elucidated, nevertheless, accumulating evidence points towards both GABAergic and glutamatergic actions of both extracts [10,17-24].
While GABA receptor binding activity is relatively well studied in anxiolytic botanicals, one of the other mechanisms that induce GABA levels to increase in the brain and potentially cause anxiety reduction is the inhibition of the enzyme GABA-T [25-27].
Anxiety-related cognitive and behavioral disorders often result from low levels of the inhibitory neurotransmitter GABA [28] and an increased glutamate release [29].
A precise combination of 120 mg VAL and 80 mg MEL standardized extracts (Songha® Night) has been shown to improve sleep quality in patients with mild to moderate sleep disturbances and to have anxiolytic effects, with no decrease in vigilance or mental performance, no additive effect with alcohol in the psychomotor functioning, and rare or inconsistent hangover symptoms, [30,31], as opposed to benzodiazepines and other sedative compounds. The neurotransmitters GABA and glutamate are both involved in the pathophysiology of sleep and mood disorders; published data point to an increase of glutamate release and a deficit of GABA [28, 29, 32-34].
Hence, we tested the hypothesis that the combined mechanism of action of the two extracts contained in Songha® Night may induce a synergistic effect modulating the levels of GABA and glutamate in the Central Nervous System (CNS). For this reason, we measured the levels of glutamate and GABA in the brain of rats exposed to chronic stress and treated with either VAL, or MEL, or a combination of both extracts. We selected the Unpredictable Chronic Mild Stress (UCMS) animal model, as most effects of UCMS can be reversed by antidepressant agents, illustrating a strong predictive validity [35].
Six to eight-week-old female Wistar rats were purchased from Charles River (Calco, Italy). Animals were left to acclimate for 1 week before entering the study. Rats were then divided in five groups (ten animals per group) and stressed using the UCMS technique. Institutional and national guide for the care and use of laboratory animals was followed (Prot. N. 15-2013).
MEL and VAL standardized extracts were supplied by Ginsana SA (Bioggio, Swizerland). The analytical markers for VAL are valerenic acids, and the fixed ratio herbal drug:dry extract = 4.5:1; while the MEL analytical marker is rosmarinic acid (> 1.8%) and the extract is standardized on the fixed ratio herbal drug:dry extract = 5:1.
L-[3,4-3H]-Glutamic acid (49.9 Ci/mmol) was obtained from Perkin Elmer Life and Analytical Sciences, Inc. LCCG-I (2S,1S,2S)-2-(Carboxycyclopropyl)glycine and L-AP4 (L-(+)-2-Amino-4-phosphonobutyric acid) were obtained from Tocris Bioscience (Ellisville, MO). All other reagents were obtained from Sigma-Aldrich Co.
The UCMS model involves the chronic sequential application of a variety of extremely mild stressors in a random order, so that they are unpredictable. The stress regimen is applied for three weeks. Different stressors are presented:
• Short periods (3 h) of food and water deprivations
• Confinement in a small tube (1 h)
• Paired housing (exposure of a rat to another stressed rat in the same cage) (2 h)
• Reversal of the dark/light cycle
• 45° tilt cage (1 h)
• Forced swim (30 min)
• Housing with damp sawdust
• Access to an empty bottle (2 h).
Rats were divided in five groups. Each group of ten animals was treated as follows:
1. Untreated and not stressed
2. Untreated and stressed
3. Treated with MEL (15 mg/kg) extract and stressed
4. Treated with VAL (20 mg/kg) extract and stressed
5. Treated with combination and stressed
Exact doses were established after preliminary experiments.
Treatment administration started one week after the beginning of the stress procedure and continued for two more weeks. After treatments, brain levels of glutamate and GABA were sampled using a microdialysis technique.
For the microdialysis experiments, rats were anesthetized with Isoflurane 1.5-2% V/V and placed in a Kopf stereotaxic frame. Intracerebral guide cannulae (OD 635 lm; ID 457 lm) provided by dialysis probe were implanted close to the lateral hippocampus and fixed to the skull. Dialysate samples were collected and frozen immediately on dry ice and were stored at -70 ℃ before analysis.
Analyses were carried out for both substances as previously described [35] by High-Performance Liquid Chromatography (HPLC) with laser-induced fluorimetric detection after o-phthalaldehyde derivatization with little changes. Briefly, an Ascentis Express C18, 100 mm × 4.6 mm, 2.7 μm column was used. The mobile phase consisted of methanol-potassium acetate (0.1 M) adjusted to pH 5.52 with glacial acetic acid. A gradient (flow rate 0.9 mL/min) of three linear steps- from 25-43% methanol (1 min), from 43-70% methanol (10 min) and from 70-90% (1 min) - followed by an isocratic hold at 90% methanol (1 min) and then back from 90-25% methanol (1 min). One volume (10 µL) of dialysate was mixed with 1 µL of the o-phthalaldehyde derivatization reagent in a glass capillary tube and injected after 1.5 min. Standard curves were linear over the concentration range of 25-1000 fmol/µL. The minimum detectable concentration was 3 fmol/µL.
GABA-T activity was determined following the method of Sherif, et al. [36]. The extracts were tested in triplicate at final concentrations in the range 0.01-1000 mg/L (in wells) in order to determine the EC50 concentration. The relative activity of each concentration was compared with the control, which was set to 100%. One unit of enzyme activity was designated to the amount of enzyme activity that converts 1.0 µmole of GABA to succinic semialdehyde and then to succinate per minute with a stoichiometric reduction of 1.0 µmole of NADP+ at pH 8.6 at 25 ℃.
Receptor binding competition assays were performed using purified synaptosomes isolated from the ventral and the dorsal hippocampus. The reaction was initiated by the addition of tissue (100 μg protein) to tubes containing 1 mM of different Ionotropic Glutamate Receptor (iGluR) and mGluR ligands and 20 nM [3H] Glutamic acid in a final volume of 500 μL of 50 mM Tris-HCl/100 mM KCl buffer, pH 7.4. The nonspecific binding was determined in the presence of 1 mM nonradioactive glutamate. All samples were incubated on ice (0-4 ℃) for 40 minutes. The assay was stopped by centrifugation for 30 min at 6,700 g; the supernatant was extracted and the pellet was washed two times with 1 mL of ice-cold buffer. The pellet was suspended in 500 μl of buffer. Radioactivity of the samples was quantified in a Beckman LS 6500 Multipurpose Scintillation Counter with 1 mL of Universal ES scintillation cocktail.
The mGluR ligands evaluated were LCCG-I (2S,1'S,2'S)-2-(Carboxycyclopropyl)glycine and L-AP4 (L-(+)-2-Amino-4-phosphonobutyric acid).
Data were analyzed using the univariate Analysis of Variance (one-way ANOVA), followed by Bonferroni t-test for multiple comparisons. Statistical significance was accepted when p < 0.05.
To assess whether VAL and MEL were able to change the levels of GABA and glutamate in animals subjected to stress, rats were stressed with the UCMS procedure.
Brain levels of GABA and glutamate before and after UCMS chronic stress are shown in Figure 1 and Figure 2 respectively. Three weeks of stress decreased the brain levels of GABA significantly (p < 0.001). The treatment with MEL or VAL increased the level of GABA in treated rats compared to untreated stressed animals, but the comparison was not statistically significant. However, the combined treatment with both extracts produced significantly higher levels of GABA than untreated stressed animals (p < 0.001) as well as to stressed animals treated with MEL (p < 0.001) or VAL (p < 0.01). The combined treatment restored the brain levels of GABA at the same levels of unstressed animals.
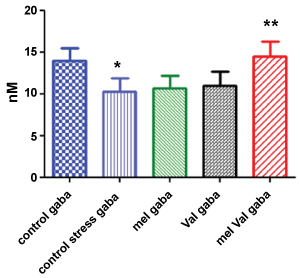 Figure 1: Brain concentration of GABA (nM) in stressed rats vs. control. Control = untreated, unstressed; Control stress = untreated, stressed; Mel = stressed, treated with M. officinalis; Val = stressed, treated with V. officinalis; Mel Val = stressed, treated with combination (*p < 0.001 vs. control; **p < 0.001 vs. control stress).
View Figure 1
Figure 1: Brain concentration of GABA (nM) in stressed rats vs. control. Control = untreated, unstressed; Control stress = untreated, stressed; Mel = stressed, treated with M. officinalis; Val = stressed, treated with V. officinalis; Mel Val = stressed, treated with combination (*p < 0.001 vs. control; **p < 0.001 vs. control stress).
View Figure 1
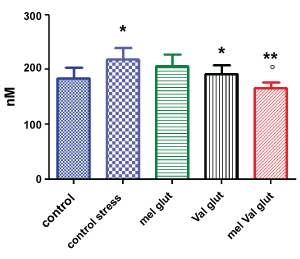 Figure 2: Brain concentration of glutamate (nM) in stressed rats vs. control. Control = untreated, unstressed; treated with M. officinalis; Val = stressed, treated with V. officinalis; Mel Val = stressed, treated with combination (* = p < 0.05 vs. control; ** = p < 0.001 vs. control stress; ºp < 0.05 versus val glut).
View Figure 2
Figure 2: Brain concentration of glutamate (nM) in stressed rats vs. control. Control = untreated, unstressed; treated with M. officinalis; Val = stressed, treated with V. officinalis; Mel Val = stressed, treated with combination (* = p < 0.05 vs. control; ** = p < 0.001 vs. control stress; ºp < 0.05 versus val glut).
View Figure 2
Regarding glutamate brain levels, induced stress increased its levels significantly (p < 0.05). The treatment of VAL alone decreased the levels of glutamate significantly (p < 0.05) compared to untreated stressed animals, while MEL alone was ineffective. Surprisingly, the combination of the two extracts significantly decreased the levels of glutamate, compared both to stressed animals (p < 0.001) and to stressed animals treated with VAL alone (p < 0.05). Also, in this case, the combined treatment restored brain levels of glutamate at levels similar to unstressed animals.
The effect of VAL and MEL extract concentrations were dose dependent in presence of [3H] glutamate, as shown in Figure 3 and Figure 4. To test the hypothesis that VAL and/or MEL may reduce the brain levels of glutamate acting at presynaptic levels, binding assays were done in presence of different types of mGluR ligands.
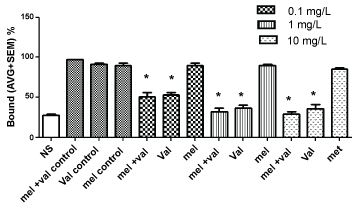 Figure 3: Interactions of V. officinalis (val), M. officinalis (mel), and its combination on metabotropic glutamate receptors (mGluR), in presence of LCGG, a Group II metabotropic glutamate receptor agonist. (*p < 0.01).
View Figure 3
Figure 3: Interactions of V. officinalis (val), M. officinalis (mel), and its combination on metabotropic glutamate receptors (mGluR), in presence of LCGG, a Group II metabotropic glutamate receptor agonist. (*p < 0.01).
View Figure 3
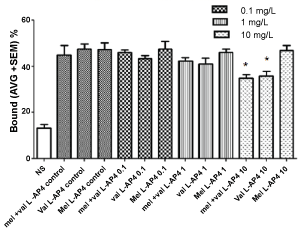 Figure 4: Interactions of V. officinalis (val), M. officinalis, and its combination on metabotropic glutamate receptors (mGluR), in presence of L-AP4, a Group II metabotropic glutamate receptor agonist. (*p < 0.01 vs. control).
View Figure 4
Figure 4: Interactions of V. officinalis (val), M. officinalis, and its combination on metabotropic glutamate receptors (mGluR), in presence of L-AP4, a Group II metabotropic glutamate receptor agonist. (*p < 0.01 vs. control).
View Figure 4
VAL extract in presence of LCCG-I or L-AP4A Group II metabotropic glutamate receptor agonists decreased [3H] glutamate binding, showing the ability to bind these receptors. No effects were observed with MEL extract alone or in combination (Figure 3 and Figure 4).
The inhibitory activity of MEL and VAL extracts on GABA-T is shown in Figure 5. The MEL extract inhibition was dose dependent and started at 5 µg/ml. At the highest dose tested, it was close to 90%. VAL extract shows about 30% inhibition at the highest concentration tested. Interestingly, the combination of MEL and VAL could inhibit GABA-T more effectively than MEL and VAL alone.
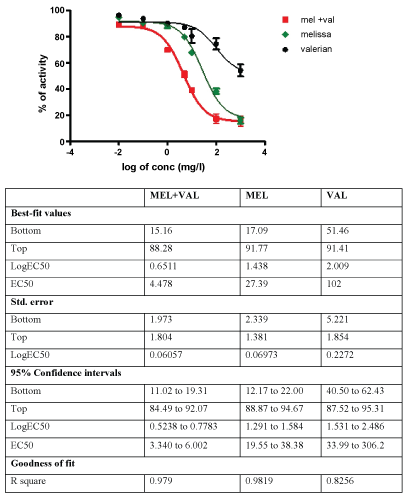 Figure 5: Inhibitory activity of different concentrations of valerian (Val), Melissa (Mel), and its combination on GABA Transaminase (GABA-T).
View Figure 5
Figure 5: Inhibitory activity of different concentrations of valerian (Val), Melissa (Mel), and its combination on GABA Transaminase (GABA-T).
View Figure 5
The amount of both neurotransmitters GABA and glutamate and sleep or mood are striclty correlated: GABA has relaxing properties whereas glutamate shows excitatory effect [32-34].
Antidepressant and mood-stabilizing treatments appear to raise cortical GABA levels and ameliorate GABA deficits in patients with mood disorders; while acute stress is associated in preclinical studies with increased excitatory amino acid transmission in areas of the forebrain. Restraint stress, tail pinch, forced swimming, foot shock, and anxiogenic drugs have been shown to increase the efflux of glutamate as measured by microdialysis in vivo [34,37-41].
The role of GABA has long been regarded as central to the modulation of anxiety and this neurotransmitter system is the target of benzodiazepines and related drugs used to treat anxiety disorders. It has been estimated that at least one-third of all CNS neurons utilize GABA as their primary neurotransmitter, and GABA ergic inhibition is essential for maintaining a balance between neuronal excitation and inhibition [42]. A number of studies have indicated that relative to healthy controls, GABA levels in plasma and cerebrospinal fluid are lower in acutely depressed patients [43-45].
The glutamatergic system is increasingly regarded as a primary mediator of psychiatric pathology and, potentially, also a final common pathway for the therapeutic action of antidepressant agents [45].
Our experiment demonstrated that the Unpredictable Chronic Mild Stress (UCMS) animal model reduces brain levels of GABA while increasing the glutamate levels. MEL and VAL show different patterns of activity: MEL and VAL administered alone could not reduce in a statistical significant way, using this model, GABA stress induced levels; however, a combination treatment did. Remarkably, upon extract combination treatment, the brain level of GABA at the end of the period of stress was close to the basal values.
Regarding the glutamate brain levels, VAL given alone counteracts the increase of glutamate induced by stress, while MEL given alone is less effective. However, the combination of VAL and MEL reduced the glutamate brain levels significantly compared to VAL given alone. This demonstrates that MEL administered alone is insufficient to counteract stress-induced increase of glutamate, but in combination with VAL can significantly counteract this effect in the brain.
Many mechanisms could be involved in the GABAergic and glutamatergic regulation exerted by the two extracts. Various studies report that VAL could stimulate the GABA transmission and inhibit GABA receptors; it may also involve benzodiazepine-like mechanism of action [10,17-20]. Kennedy, et al. found that an acute administration of 600 mg of MEL extract increased calmness and reduced alertness [24]. This pattern of mood modulation suggests the involvement of the GABAergic system in the MEL anxiolytic effects. A survey of ten anxiolytic botanicals was reported recently, and an extract of MEL was found to be the best inhibitor of in vitro GABA-T activity from rat brain [27]. Similarly, Awad, et al. [46] reported that MEL inhibits GABA-T, attributing this effect to rosmarinic acid, a major constituent of lemon balm. In our study, we confirmed that MEL is an inhibitor of GABA-T activity; however, this mechanism could be not sufficient to produce a significant effect in counteracting the stress-induced reduction of GABA levels.
With respect to the release of glutamate, we tested the hypothesis that the extracts in the study could act as agonists on mGluRs. In recent years, several studies have investigated the effects of VAL in the excitatory glutamate-mediated neurotransmission and showed that the extract of VAL and in particular valerenic acid are able to interact with Group I and Group II mGluRs, thus having calming and relaxing effects that may represent the main mechanism of action to explain the anxiolytic properties of this plant [21,22,47,48].
In our study, VAL extract in presence of LCCG-I, a Group II metabotropic glutamate receptor agonist, shows a marked decrease in the binding at all tested concentrations. This effect was seen with L-AP4 at high concentration only. Our results confirm the hypothesis of VAL interaction with the presynaptic mGluRs, suggesting a possible mechanism by which VAL extracts counteract the excess of glutamate release induced by stress. On the other hand, the extract of MEL seems to have no detectable effect on these glutamate receptors.
Our results demonstrate that the combined mechanism of action of the two extracts contained in Songha® Night may induce a synergistic effect acting on two targets, inducing the normalization of the hyperactive excitatory brain system seen during stressful conditions: MEL-inhibits GABA-T activity and increases the levels of GABA, and VAL acts on Group II mGluRs and decreases the release of glutamate. Furthermore, the two extracts together exert a unique functional synergism leading to normalization of brain levels of GABA and glutamate altered by stress. This may explain the clinical effect of the two compounds on the reduction of anxiety and the consequent relief of sleep disturbances as reported in clinical studies performed with Songha® Night.
However, the exact mechanism involved in this action is complex and not yet fully understood and it will be the subject of future investigations.
We wish to confirm that there has been no significant financial support for this work that could have influenced its outcome. Andrea Zangara has been a consultant to Ginsana SA.